Esophageal injuries of different severities may be manifested in patients following radiotherapy for thoracic, and some head and neck, cancers, and such injuries include both acute and chronic effects due to the range of molecular reactions [27,28,29]. In severe cases, the fear and pain of eating and drinking due to esophageal injury can result in cachexia, which may lead to the interruption or cessation of clinical treatment [6]. Esophageal injury greatly prolongs therapy and greatly reduces the effectiveness of radiotherapy [9]. ARIE usually appears as early as the first week of radiotherapy, and in the majority of cases, it becomes symptomatic within the first two months after radiotherapy [30]. Although numerous studies have been conducted to combat ARIE, effective and ideal treatments are still unavailable [31]. Therefore, establishing a standard and clear animal model to facilitate the prevention and treatment of ARIE is highly valuable.
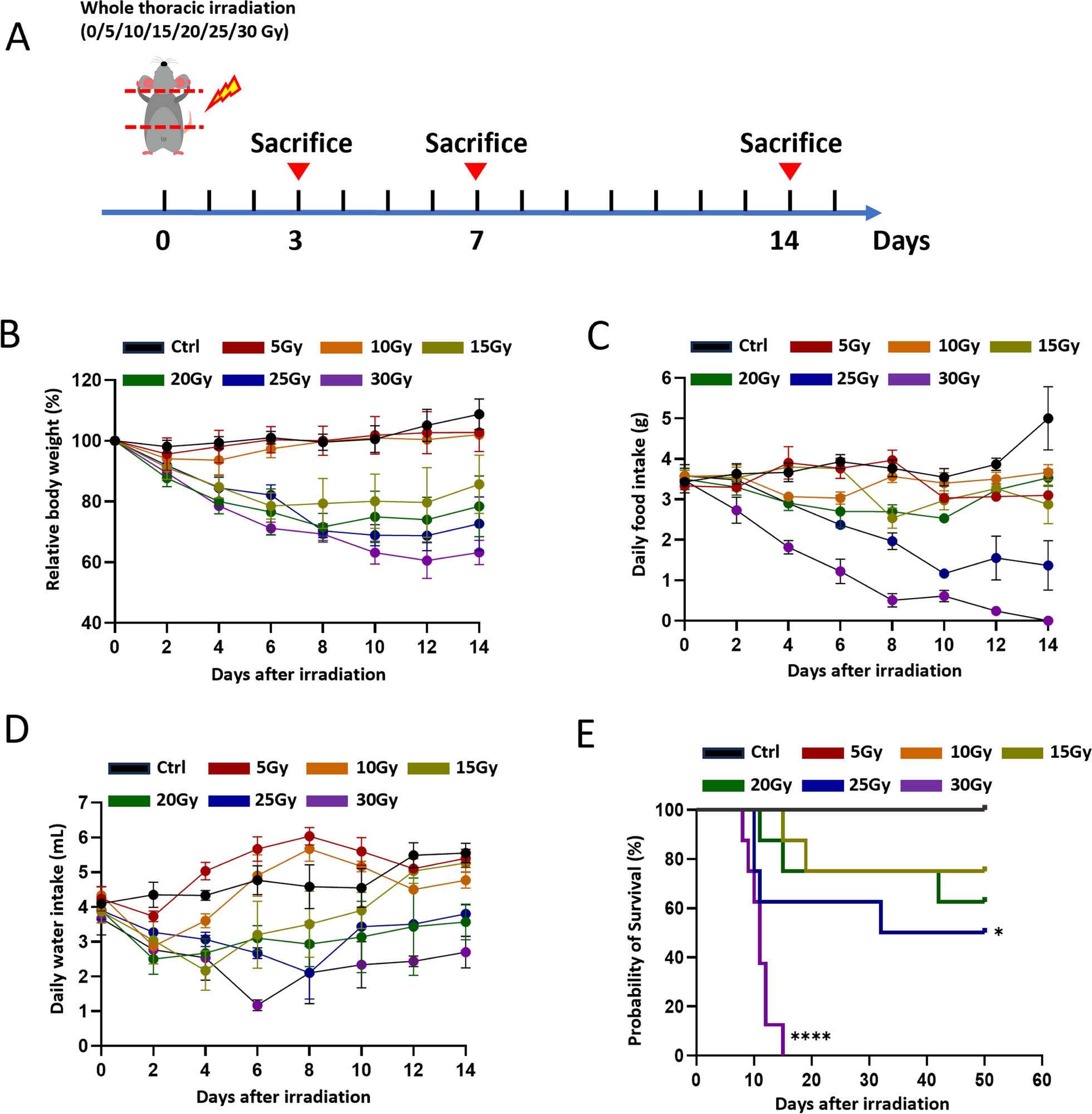
Mice are the most widely used experimental model, and many well-established genetic resources are available. In this study, we compared X-ray irradiation-induced acute esophagitis in C57BL/6 mice via a wide range of single doses. We have chosen 6 to 8-week-old male C57BL/6 mice for this study in order to better simulate the clinical scenario of radiation esophagitis in human esophageal cancer, which is more prevalent in males [32, 33]. Additionally, male mice were selected to avoid the potential interference of cyclical hormonal changes in females, which could affect the pathological processes of radiation-induced damage [34, 35]. This choice also enhances the accuracy and reproducibility of experimental data. The age of 6 to 8 weeks was selected because the organs of the mice have gradually matured and they are in good overall health, making them suitable for most experimental studies [36]. The severity of ARIE was shown to be positively dose dependent, which was like that of radiation damage in other parts of the body [37, 38]. A higher irradiation dose caused more impairments, such as worse mental status and locomotor ability, reduced body weight, and decreased water and food intake. All these symptoms are closely related to more severe esophageal injuries [39]. ARIE is characterized by a marked reduction in the thickness of the esophageal epithelium [40]. H&E staining revealed obvious epithelial damage to the esophagus of mice receiving higher doses (Fig. 2A), and this study revealed that esophageal tissues irradiated with 15 Gy or more exhibited observable degrees of radiation damage. The esophageal epithelium in the 30 Gy group was markedly thinned, with extensive necrosis, detachment, basal cell hyperplasia, and infiltration of immune cells. However, increased incidence of esophageal injuries might result in increased mortality. Therefore, it is necessary to determine an optimal radiation dose for laboratory use. A comparison of ARIE caused by different doses revealed that a single 20-Gy dose of thoracic irradiation could successfully induce ARIE while ensuring a certain survival rate; thus, we propose that 20 Gy of thoracic irradiation as the most appropriate dose for establishing an ARIE model in mice.
The progression of typical radiation injury involves five sequential stages: the injury initiation stage, the signal transduction stage, the signal amplification stage, the ulcer formation stage, and the healing and repair stage [41, 42]. Here, we focused on two of these stages, the initiation phase and the healing and repair phase of ARIE after 20 Gy of irradiation. Severe epithelial injury and immune cell infiltration were observed on Day 7. There was a significantly thinned esophageal epithelium, discontinuity of the epithelial layer, and disordered epithelial cells, which were coupled with a substantial loss of CK14-positive basal layer cellularity (Fig. 3). DNA damage and ROS production are common pathological alterations associated with radiation injury. This was further characterized by a decrease in the number of PCNA- and PHH3-positive basal layer cells, an increase in the number of γH2AX-positive cells, the increasing of ROS signals- and TUNEL-positive epithelial cells (Fig. 4). We demonstrated that esophageal tissues had obvious inflammatory responses on Day 7. The results revealed a significant increase in the number of neutrophils, T cells, and macrophages (Fig. 6A, B), as well as increased gene expression levels of proinflammatory factors (IL-1β, IL-6, and TNF-α) and decreased expression of the anti-inflammatory factor IL-10 (Fig. 6C–F). Inflammation disrupts the homeostasis of cellular proliferation and differentiation and leads to subsequent barrier disruption and long-term mucosal damage. These results suggest that the pathological mechanisms of radiation damage may involve the interplay of DNA damage, inhibition of cell proliferation, and ROS and inflammatory responses. DNA damage is one of the core mechanisms of radiation injury. The increase in γH2AX-positive cells indicates the formation of DNA double-strand breaks, while the decrease in PCNA- and PHH3-positive cells reflects the reduced capacity for cell proliferation. This may be due to DNA damage activating cell cycle checkpoints, which leads to cell cycle arrest and subsequently inhibits normal cell proliferation [43]. Additionally, the increase in ROS further exacerbates cellular damage. ROS can directly attack cell membranes, proteins, and DNA, disrupting cell structure and function, and can also trigger apoptosis and inflammatory responses by activating various signaling pathways [44, 45]. Finally, although the infiltration of inflammatory cells and the release of pro-inflammatory factors help to clear damaged cells and tissue debris, creating favorable conditions for tissue repair, excessive inflammatory responses can further exacerbate tissue damage [46]. In this study, the intense inflammatory response observed on Day 7 may be one of the key factors leading to severe esophageal epithelial damage.
In addition, esophageal injury in the 20 Gy group presented clear signs of regeneration and healing on Day 14, characterized by a notable increase in epithelial thickness, a diminished inflammatory response, and well-aligned basal cells. Moreover, both the number of inflammatory cells and the number of inflammatory factors were significantly lower at this time than on Day 7. On day 14, the increased thickness of the esophageal epithelium and the well-aligned basal cells, along with the recovery of CK14-positive stem cells, indicate that the esophageal tissue possesses a certain capacity for self-repair, which may be associated with the activation and proliferation of stem cells. Meanwhile, the significant reduction in the number of inflammatory cells and inflammatory factors, and the attenuation of the inflammatory response, provide a favorable microenvironment for tissue repair and reduce further damage to the tissue caused by inflammation [46]. Therefore, the most severe esophageal injury associated with ARIE occurred on Day 7, and the mice recovered on Day 14 after 20 Gy of irradiation, unlike other radiation-sensitive tissues, such as the small intestine and bone marrow [47, 48]. This difference may be related to the characteristics of the stem cells and the microenvironment of the esophageal tissue.
However, it can be observed from Fig. 3A-D that the epithelium, stratum corneum, and pathological grading decreased again after irradiation for 14 days. Studies have shown that the process of esophageal epithelial thickening followed by a return to normal thickness involves mechanisms such as cell proliferation, differentiation, apoptosis regulation, and remodeling of the extracellular matrix (ECM) [49,50,51,52,53,54]. Firstly, it may be related to the decrease in proliferating cells. In the early stage of injury repair, basal layer stem cells proliferate massively to fill the damaged area, resulting in epithelial thickening. As repair progresses and the epithelial structure becomes intact, proliferation signals weaken, proliferation slows down, and excess cells are gradually reduced through normal metabolism [50]. Secondly, it may be related to the differentiation and maturation of cells. New epithelial cells differentiate into mature keratinocytes, forming a normal stratified squamous epithelium. The connections and functions between cells are perfected, stabilizing the epithelial thickness [50]. Additionally, it may be associated with apoptosis. During the esophageal epithelial regeneration, apoptosis is strictly regulated. The HGF/c-MET and NGF/TrkA signaling pathways inhibit apoptosis, promoting the survival and regeneration of esophageal epithelial cells [49,50,51]. As repair is completed, the activity of these pathways decreases, apoptosis returns to normal levels, and excess cells are cleared through apoptosis, restoring epithelial thickness to normal [49,50,51]. Moreover, it may be associated with the remodeling of the extracellular matrix. ECM components such as collagen and fibronectin increased during the repair process, providing support for new epithelial cells. After that, ECM components are remodeled, excess collagen is degraded, and the matrix returns to its normal structure, restoring epithelial thickness to normal [52]. Furthermore, it may be associated with the resolution of the inflammatory response. After repair is completed, inflammatory signaling pathways are downregulated, inflammatory cells decrease, and the inflammatory response subsides. The stimulation of cell proliferation and matrix synthesis by inflammatory factors is reduced, leading to the stabilization of epithelial thickness [53]. Lastly, it may be related to the regulation of quiescent stem cells. Studies have found that a population of quiescent stem cells (QBCs) exists in the basal layer of the esophagus. These cells are activated to participate in repair after injury. After repair is completed, QBCs re-enter a quiescent state, reducing excessive cell proliferation and aiding in the recovery of epithelial thickness [54]. In summary, the return of esophageal epithelial thickness to normal levels after injury repair is the result of the combined action of cell proliferation and apoptosis balance, ECM remodeling, and inflammatory response resolution.
Presently, the construction of animal models for ARIE predominantly utilizes rats, mice, and rabbits as experimental subjects. The prevailing experimental paradigm in the majority of studies involves administering a single high-dose irradiation regimen, while a few have attempted low-dose fractionated irradiation [8,9,10, 13]. Damage assessment mainly relies on histopathological observations of esophageal epithelial necrosis, inflammatory cell infiltration, and the expression levels of inflammatory factors [11, 12, 55]. Esophageal acute radiation injury typically manifests between the 5th and 14th day after irradiation, while the recovery time remains unclear. In this study, we used single-dose thoracic X-ray irradiation in C57BL/6 mice to construct the model. The extent of esophageal epithelial damage was subsequently evaluated through histopathological analysis and quantification of inflammatory cytokine expression levels. In contrast to prior studies, the current research not only thoroughly elucidated the roles of DNA damage, inhibition of cell proliferation, ROS production, and inflammatory response in the injury and repair processes, but also preliminarily clarified the repair mechanisms, emphasizing the key role of stem cell activation in tissue repair. Through a comparative analysis of various irradiation doses, we established that 20 Gy represents the optimal irradiation dose, offering a standardized reference model for future studies. Additionally, this study clarifies the process of esophageal radiation injury and repair, which has certain implications for dose optimization in the radiotherapy of esophageal cancer. In clinical treatment, it is necessary to balance the relationship between radiotherapy dose and tissue injury to avoid severe esophageal damage caused by excessively high doses. At the same time, in the radiotherapy of esophageal cancer, the radiotherapy dose is closely related to the therapeutic effect. Studies have shown that appropriately increasing the radiotherapy dose can improve local–regional control, thereby enhancing overall survival (OS) and progression-free survival (PFS) in patients [56]. However, beyond a certain dose, radiation toxicity can significantly increase and even offset survival benefits. Therefore, determining the optimal radiotherapy dose is crucial for improving the efficacy of esophageal cancer radiotherapy.
Despite these interesting findings, there are still shortcomings in the current study. First, the model we established was relatively simple and did not explore the potential mechanisms of radiation damage and regeneration of the esophagus in depth, which will be further investigated in future studies. Second, we adopted only a single dose of X-ray irradiation for the purpose of operational convenience in establishing an experimental mouse model. Since fractioned irradiation is used in clinical practice, such an irradiation strategy will also be tested in the future. Finally, we used whole thoracic irradiation which may also damage some vital organs, such as the lung and heart, which can not exactly mimic clinical circumstances. Although this study mainly focuses on acute injury and repair, long-term fibrotic damage is also an issue that needs attention in the radiotherapy of esophageal cancer. Long-term fibrosis can lead to serious complications such as esophageal stricture, affecting patients' quality of life. In clinical practice, long-term follow-up of esophageal cancer patients receiving radiotherapy is necessary to monitor potential late complications and take timely intervention measures. Therefore, future research needs to further explore strategies for preventing and mitigating fibrosis.




Comments (0)