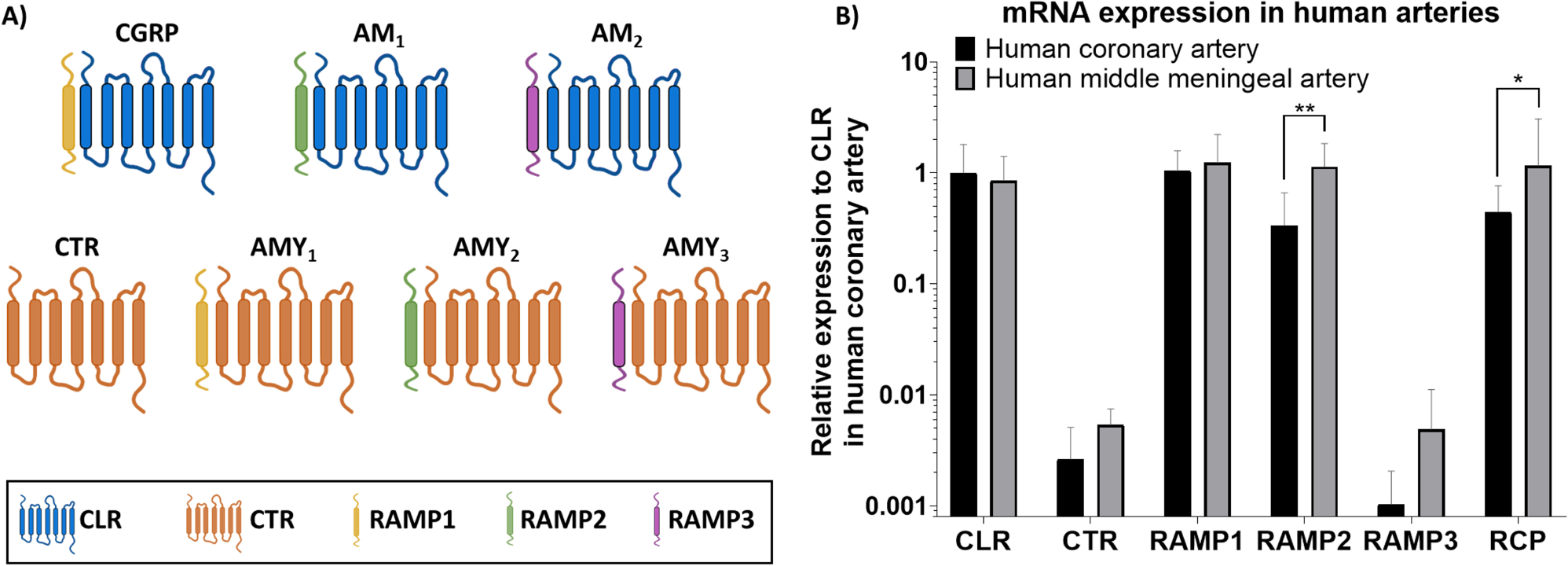
The current study shows that the expression of the canonical CGRP receptor (CLR-RAMP1) is similar in human coronary arteries and human middle meningeal arteries, while the expression of the adrenomedullin 1 receptor (CLR-RAMP2) differs, with a higher expression in human middle meningeal arteries. The current expression data matches largely with the expression as reported in the Human Protein Atlas [16] for smooth muscle tissue normalized expression (nTPM), with the highest expression of β-actin, followed by GAPDH, RAMP1 and RAMP2, which is exactly the order of expression in human middle meningeal arteries and similar to human coronary arteries except for the lower expression of RAMP2 in this tissue. Indeed, CTR expression is reported to be absent or low in this tissue, again matching our data. It should be noted that our isolated arteries do not exclusively consist of smooth muscle tissue, but also connective tissue and endothelial cells. However, the arteries consist of a thick layer of smooth muscle tissue and only a monolayer of endothelial cells, suggesting that the main contribution to the RNA content in our homogenized arteries is from smooth muscle cells. Further research is needed to determine the relation between mRNA expression and protein expression of all receptor subunits in the vasculature. However, due to the scarcity of the human arteries and the limited amount of tissue obtained per patient, it was not possible to assess both protein expression and mRNA expression within the same tissue. In line with this, mRNA expression and functional responses could not be measured in tissue of the same donor. Therefore, we cannot draw conclusions on whether the functional responses match the expression data in an individual donor. Furthermore, we only have information from the total homogenized arteries and cannot separate the results per cell type (i.e. endothelial cells or smooth muscle cells), or per subcellular compartment. Further research is necessary to determine where exactly the receptor subunits are expressed, and what the physiological relevance of this expression is likely to be.
The findings of the current study in coronary arteries are in accordance with a previous studies in both human and porcine coronary arteries in which mRNA of subunits of the canonical CGRP receptor and the adrenomedullin 1 receptor was detected, while CTR and RAMP3 could not be detected porcine coronary arteries [17, 18]. In addition, RAMP1, RAMP2, RAMP3 and CLR expression have been previously detected in human middle meningeal arteries [19]. The current study offers a direct comparison between these two vascular tissues and allows quantification of expression of the different subunits. Interestingly, in contrast to the vasculature, expression of CTR was previously detected in the human trigeminal ganglion and dorsal root ganglion, where it is likely involved in pain transmission [20, 21].
Interestingly, in depth analysis of the expression data shows two types of expression patterns in human middle meningeal arteries, with some patients having higher RAMP1 expression and others having higher RAMP2 expression. Moreover, our functional results in human middle meningeal arteries show the same two patient groups, of which some respond clearly to the adrenomedullin receptor antagonist AM22-52, while others do not. AM22-52 is the truncated version of adrenomedullin and is an antagonist at the adrenomedullin 1 receptor, while at a lower potency it can also antagonize the adrenomedullin 2 receptor [22]. The fact that both our functional results and the mRNA expression data can be divided in two populations could suggest that the mRNA expression of RAMP1 and RAMP2 reflects the protein expression in these tissues. In contrast, the adrenomedullin 1 receptor antagonist AM22-52 had no effect in human coronary arteries, in which RAMP1 expression is predominant. The absence of antagonistic effects of AM22-52 in coronary arteries has been described previously [17, 18].
The novel anti-migraine drugs targeting the canonical CGRP receptor (i.e. gepants and the monoclonal antibody erenumab) are designed to target the hydrophobic pocket between CLR and RAMP1, thereby preventing direct binding of CGRP to the receptor [23, 24]. Gepants and erenumab have a high affinity for the CGRP receptor, and no or only low affinity for the adrenomedullin 1 receptor (CLR-RAMP2) or the adrenomedullin 2 receptor (CLR-RAMP3) [22, 25,26,27,28]. Therefore, treatment response could be affected by the exact receptor expression in HMMA, which could serve as a direct target for antimigraine medication, and as a proxy for what is happening in the trigeminovascular system, an important structure involved in the pathophysiology of migraine [29]. Interestingly, the observations in the current study showing two types of expression patterns of RAMP1 and RAMP2 in HMMA donors, could suggest that also two types of (migraine) patients exist, who possibly respond differently to treatment. If in some patients the RAMP2 expression in the human middle meningeal artery is higher than RAMP1, CGRP signalling could continue via the adrenomedullin 1 receptor even if the CGRP receptor is blocked, resulting in less effective treatment with the CGRP receptor antagonists. Future studies should determine whether receptor subunit expression differs between migraine patients and healthy controls. Unfortunately, the migraine status of the patients included in the current study, or their medication use, is unknown because of ethical regulations, anonymizing our donors.
One of the interesting findings of the current study is the differential expression of RCP, with higher expression in the human middle meningeal arteries compared to human coronary arteries. The exact role of this receptor subunit has been debated [8]. RCP is located on the intracellular side of the G protein coupled receptor CLR, and was shown to co-immunoprecipitate with both the canonical CGRP receptor and the adrenomedullin 1 receptor [30]. Therefore, since both receptors are abundant in human middle meningeal arteries, while the canonical CGRP receptor is predominant in human coronary arteries, the increased expression of RCP in human middle meningeal arteries could be because it forms a functional unit of both receptors present in the meningeal vasculature. It has previously been shown that loss of RCP does not affect receptor density or receptor binding, while it does affect intracellular cAMP production [30, 31], suggesting a role for intracellular signalling. Interestingly, depletion of RCP had a larger effect on cAMP levels after stimulation with CGRP compared with stimulation with adrenomedullin [30]. It should be noted that RCP protein levels were poorly correlated with RCP mRNA expression in mouse uterus during pregnancy [32], and it is suggested that RCP mRNA levels might not accurately predict RCP function.






Comments (0)