
Remember me





Male Sprague–Dawley rats weighing 50 to 250 g (Charles River, L’Arbresle, France) were housed at 22.6 ± 1 ̊C in plastic cages (size: 425 × 266x185 mm; 2–3 rats per cage) on soft bedding with ad libitum water and food pellets under an inverted 12/12 h light/dark cycle for at least 1 week before the experiments. Every effort was made to minimize the number of animals used: numbers of animals were selected according to previous experience [7, 12], i.e. with a trade-off between reaching routine sample sizes for each type of experiments and minimizing numbers of animals for pain experiments. Experiments were performed on 86 animals (Fig. 1A). All experiments, analysis, and reporting were ARRIVE-compliant (Animals in research: reporting in vivo experiments). Animal experiments were performed according to the ethical guidelines set by the International Association for the Study of Pain [22] and European Directive 2010/63/EU on the protection of animals used for scientific purpose. The protocols applied here for animal care and use were approved by the Clermont Auvergne University Institutional Review Board and authorized by the French Ministry of Primary, Secondary and Higher Education, and Research (n ̊ CE 27–12, 26–12, 700.02).
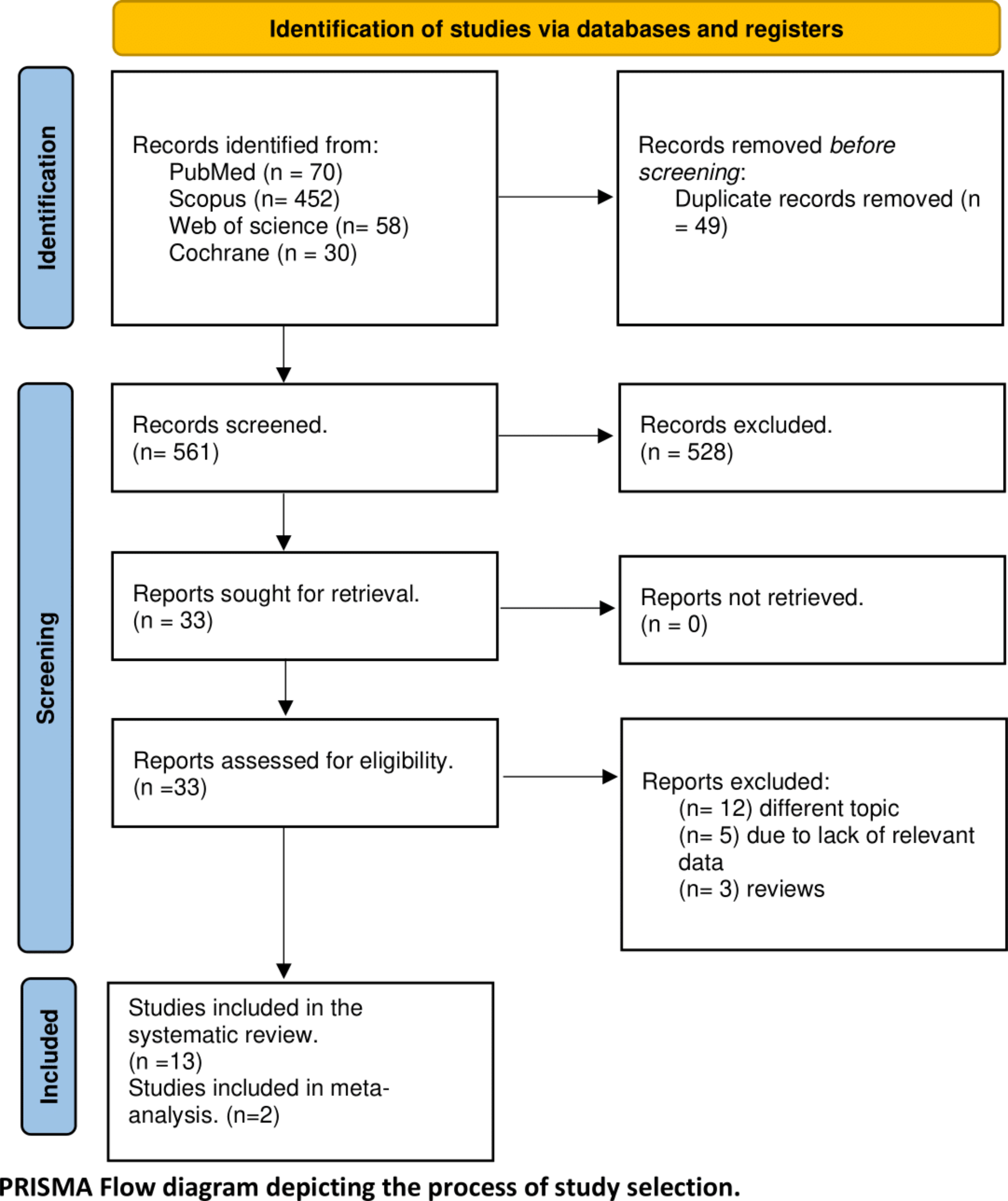
Fig. 1
Flow chart of animal experiments and schematic representation of the experimental design. A Number of animals used in each experiment: (1) animals lost during the surgery of cannula placement or blocked cannula; (2) animals excluded during electrophysiological recordings because no neurons were recorded (animals lost during surgery, loss of signal, neurons not found); (3) animals excluded because the injection site of aCSF or propranolol were outside the LC. B Rats were acclimated to handling and sensory testing 7 days before and after the surgical implantation of a dural cannula (performed on D -7). In the migraine model group, rats received four injections of inflammatory soup (IS). Mechanical sensitivity of ophthalmic cutaneous territory was tested every day, one week after the surgery. For the combined behavioral/electrophysiological study, animals received, in addition to dural injection, a daily intra-LC microinjection of propranolol or aCSF. Electrophysiological recordings were performed either in naive (no surgery, no injections) or sensitized rats, in which case recordings were realised either one or two days after the fourth IS injection (IS4 in the rest of the manuscript)
Experimental designThe study is divided in three parts. First, to investigate LC/TCC physiological relationship, double-site electrophysiology was performed in naive rats. Artificial cerebrospinal fluid (aCSF) was microinjected in the LC during recordings as a control condition for pharmacological manipulations; similar recordings were conducted in a rat model for the recurrent activation of dural nociceptors to assess potential changes. Second, to study the effects of propranolol on TCC sensitization, behavioral effects of propranolol microinjections in LC were assessed in the model. The same rats were subsequently subjected to electrophysiological recordings. Finally, to identify the preventive action of propranolol microinjection into the LC, RNAScope fluorescent in situ hybridization, in vivo double-site and whole-cell patch-clamp electrophysiological recordings were conducted in naive animals.
Dural-evoked trigeminal activationAs previously described [7, 12], rats underwent surgery after one week of habituation to the experimenter and the observation room. After anaesthesia, a cannula was carefully inserted (1 mm anteroposterior and mediolateral from bregma) into a 0.5 mm diameter craniotomy, fixed to anchor screws with dental cement, and the skin was sutured. After surgery, rats were housed separately and recovered for at least one week before sensory testing while their state was closely monitored. Injections (10 μl) of IS or aCSF were performed under brief anaesthesia (3% isoflurane; ≤ 3min) through the intracranial cannula, four times, at 2 to 3-day intervals during the 8-day injection protocol (Fig. 1B).
Intra-LC injectionsDuring the surgery to implant the dural cannula, a second stainless-steel guide cannula (o.d. 0.6mm, i.d. 0.5mm) was implanted unilaterally in the brainstem, aimed at the LC ipsilateral to the first cannula (anteroposterior, –9.68 mm; mediolateral, 1.4 mm from bregma; dorsoventral, 6 mm), and fixed with the screws and dental acrylic used for the dural cannula. The post-surgery monitoring is the same as described above. Animals for the behavioral study received daily microinjections (250 nL) of aCSF or propranolol into the LC. The microinjections were performed under brief anesthesia (3% Isoflurane; ≤ 3min) using a needle (26 G) introduced through the guide cannula until its lower end extended 1mm below the guide cannula. The needle was left in place for one additional minute after the injection.
The days of concomitant administration of IS or aCSF on the dura, animals were anesthetized a single time, and microinjection was carried out 1 min before dural administration of aCSF or IS.
After each experiment, the location of the microinjection was confirmed by cryosectioning the brainstem. When the site was located outside the LC, the rat was excluded from the analysis.
Behavioral testingAs described previously [7], the week before surgery, rats were acclimated to handling, to the behavioral context and to innocuous mechanical stimulation (sub-threshold to elicit the appropriate behavioral response). Mechanical cutaneous sensitivity of the face was then assessed before and after surgery to ensure the absence of surgery-induced sensitization, and every day during the 8-day injection protocol, on both IS injection and interictal days. Face static mechanical threshold were measured by applying von Frey (VF) hairs (Bioseb, France) to the midline of the forehead. For cephalic mechanical dynamic sensitivity, responses to gentle air-puff stimuli on the face were scored (dynamic mechanical scores: DMS) according to Vos et al. [23].
In vivo extracellular recordingsAfter the animals were anesthetized in a box with 3% isoflurane in a N2O/O2 mixture (2:1), the trachea was cannulated and the carotid artery and external jugular vein catheterized. Animals were then paralyzed by an intravenous perfusion of vecuronium bromide and artificially ventilated with a volume-controlled pump. Levels of isoflurane, O2, N2O and end-tidal CO2 were measured by an anaesthetic gas analyser (Drager Vamos) during the entire experimental period. The arterial catheter was attached to a calibrated pressure transducer connected to an amplifier for continuous monitoring of the mean arterial blood pressure. The analog output from the blood pressure amplifier was connected to a computer data sampling system (Cambridge Electronic Design 1401plus interface, CED; Cambridge, UK). The animals were placed in a stereotaxic frame with the head fixed in a slightly ventroflexed position. A craniotomy was performed to allow chemical and electrical stimulation of the dura and passage of the electrode for LC recordings. After surgery, the level of isoflurane was progressively reduced to 1.2% in pure O2 and maintained at this level during the recording period.
Unitary extracellular recordings were made from the right TCC with glass micropipettes (0.5–2 MΩ) filled with a mixture of 5% NaCl and pontamine sky blue. TCC electrophysiology was conducted as previously described [7]. Briefly, wide-dynamic range (WDR) neurons with both cutaneous and meningeal receptive fields were recognised based on their responses to cutaneous mechanical non-noxious (brushing with a soft brush) and noxious (pinch with forceps) stimulations of their cutaneous receptive fields and to meningeal electrical stimuli delivered through small silver balls.
Recordings were simultaneously made from the ipsilateral LC with a tungsten microelectrode (0.5 MΩ, World Precision Instrument, USA), glued to a glass micropipette for microinjections. Because a rectangular insertion of the recording electrode into the LC would affect the most sensitive part of the meninges and bear a great risk of haemorrhage, the microdrive was tilted to a 30° angle from the horizontal plane, and more anterior coordinates were used (-5.7mm posterior and 0.7mm lateral from bregma). A meningeal incision was made parallel to the transverse sinus at the appropriate coordinates. The electrode was then slowly lowered (6.5–8.2mm) to find the LC. Some elements helped correcting coordinates, namely the mesencephalic trigeminal nucleus neurons, lateral to the LC and responding to jaw opening, as well as tail movements-responsive neurons encountered along the descent (putative cerebellar neurons). LC neurons were identified according to usually described characteristics: tonic activity, biphasic response to noxious stimuli (activation followed by inhibition), and location was later confirmed by a lesion made by passing current through the electrode at the end of the recordings.
LC multi-/single-units, and field potential recordings were both acquired through a Plexon system (on an unfiltered channel for LFP) and the above-described CED 1401plus interface/Spike2 system.
Electrical stimulationsTwo modalities of electrical stimuli, delivered either to the face (periorbital cutaneous zone) or the meninges, were used: stimuli of 0.8 ms applied every 1.5 s (traditionally used for TCC recordings) or stimuli of 5 ms applied every 2 s (that were previously described as more suitable to see some responses in the LC [24]). The face was stimulated at three times the threshold (in mA) necessary to elicit a C-fibre response in the TCC (0.8 ms stimuli). Given the impossibility to safely map the meningeal receptive field of the TCC WDR because of the near LC electrode, and therefore determine its threshold, intensity of stimulation for meningeal stimuli was kept constant across conditions and animals at 4-5mA.
Mechanical stimulationsIn all double-site recordings, mechanically evoked LC activity was probed by strong but brief stimulations of the paw and the face. Therefore, we used fine forceps (3 mm apart) to apply mild pressure to the contralateral ankle, or the extremities on the side of the ipsilateral muzzle for a duration of 0.5 s.
For the behavioral/electrophysiological study, we used two types of mechanical stimulations: (i) brushing with a soft brush (0.5-s brush stroke at 0.5 Hz during 20 s) and (ii) pressure applied with VF hairs (0.16, 0.4, 0.6, 1, 1.4, 2, 4, 6 and 8 g), each VF hair being applied once for 5 s in random order, to the most sensitive portion of the cutaneous receptive field.
Dural-evoked trigeminal activationThe exposed dura was bathed for 5 min in IS as described before [7].
Microinjection (LC)Propranolol or aCSF microinjections (250 nl) into the LC was carried over 1 min out by applying short pulses of high-pressure air with a custom-made apparatus.
Patch-clamp electrophysiological recordingsRats were deeply anaesthetized with an intraperitoneal (i.p.) overdose of chloral hydrate (7%) and decapitated. The brains were quickly removed and immediately chilled in ice-cold cutting-based saline solution bubbled with carbogen (95% O2, 5% CO2) and containing the following (in mM): 2 KCl, 0.5 CaCl2, 7 MgCl2, 1.15 NaH2PO4, 26 NaHCO3, 11 glucose, and 205 sucrose. After removal of the dura mater, the brainstem including LC was transversally sliced (350 μm thick) with a vibratome (VT1200 S, Leica Microsystèmes SAS, France). After cutting, slices were incubated at 37 °C in aCSF containing (in mM): 130 NaCl, 3 KCl, 2.5 CaCl2, 1.3 MgSO4, 0.6 NaH2PO4, 25 NaHCO3, 10 glucose (pH 7.4) bubbled with 95% O2 and 5% CO2, for a 45 min recovery period. Slices were then transferred into the recording chamber.
LC neurons were visualized using an upright microscope fitted with fluorescence optics (AxioExaminer, Carl Zeiss, Germany) and linked to a digital camera QImaging Exi Aqua (Czech Republic). Patch pipettes (5–7 MΩ resistance), made from borosilicate glass (1.5 mm O.D; PG150T-15; Harvard Apparatus, UK) were filled with an internal solution containing (in mM): 135 KCl, 0.5 mM CaCl2, 2 MgCl2, 5 KCl, 5 EGTA 5 Hepes, 5 ATP-Na2, 0.5 GTP-Na2, neurobiotin (0.05%, Vector Laboratories, Burlingame, CA, USA), dextran tetramethylrhodamine (10,000 MW, fluoro-ruby, 0.01%, Life technologies, Saint Aubin, France), pH adjusted to 7.4 and osmolarity of 290–300 mOsm.
Whole-cell voltage-clamp recordings were made at room temperature (22—24°C). Acquisitions were performed using Clampex 10 software (Molecular Devices, Sunnyvale, CA, USA) connected to a Multiclamp 700B amplifier (Molecular Devices, Sunnyvale, CA, USA) via a Digidata 1440A digitizer (Molecular Devices, Sunnyvale, CA, USA). Voltage-clamp data were low pass filtered at 2 kHz and digitized at 10 kHz. Series resistance was monitored at the beginning and end of each recording session, and data were rejected if values changed by > 20%.
At the end of the recordings, epifluorescence was used to ensure that the recorded cells were filled with dextran tetramethylrhodamine. Immunolabeling neurobiotin was carried out to check that the recorded neurons were in LC region. Slices were transferred into 4% paraformaldehyde in 0.1M phosphate-buffered solution (pH 7.4) and stored overnight at 4°C. Next, fixed slices were washed with 0.05 M Tris-buffered saline (TBS), and were incubated with Avidin DCS-rhodamine (1:200, Ref. A-2012; Vector Laboratories, Burlingame, CA, USA) for 4 h at room temperature. Subsequently, all slices were mounted on gelatinized slides in a DPX mounting medium, cover-slipped and conserved at 4°C.
RNAScope fluorescent in situ hybridizationRNAscope kit was purchased from Bio-Techne. Rats were anesthetized, perfused transcardially with 100–200 mL of PBS followed by 500 mL of 4% RNase free paraformaldehyde. The brains were dissected and post-fixed for 24h following by cryoprotection in sucrose (gradient concentration of 10%, 20%, 30%) at 4˚C. The brains were embedded in Tissue-tek and stored at -80°C, until further processing. Frozen tissue was cut on a cryostat at 15 μm, collected on slides, and processed per the manufacturer’s protocol. Every 5th section of LC was analyzed using Zeiss AxioImager M2 microscope and the software Zen 2.3 lite (Zeiss). The probes used were targeted against the rat genes for adra2a, adrb1 and adrb2.
Drugs and substancesIS consisted of 2 mM of histamine, serotonin, bradykinin, and 0.2 mM of PGE2. Inflammatory mediators were diluted in 10 mM Hepes buffer at pH 5.0 (Sigma-Aldrich) [7, 8]. Propranolol (Sigma-Aldrich) was diluted in aCSF to a concentration of 1 mg.mL−1. Vecuronium bromide (Sigma-Aldrich) was diluted in physiological serum (9 g NaCl.mL−1 H2O), filtered, and stored in 10 mL tubes at -20°C.
AnalysisSpikes extraction was performed using the built-in Spike2 tools for single- and/or multi-units, and all data were exported from Spike2 to MATLAB (R2014b) for further analysis. Unless otherwise stated, common electrophysiological analyses were performed using custom-made scripts and built-in MATLAB functions; we also used some functions from the FMA-Toolbox (http://fmatoolbox.sourceforge.net/).
Single-unit versus multi-unit activityWhile for the TCC, we analysed single-unit (SU) activity, results presented for the LC were extracted from multi-unit responses (MUA) unless otherwise stated, for several reasons: (i) single-unit activity (SUA) responses are usually quite low (that is, a cell rarely fires more than 1 or 2 spikes per electrical stimulus), requiring repetitions, but MUA yields the same results with less “noise” (thanks to the uniformity of response of a local population of cells); (ii) the modality of microinjection, first designed for behavioral studies, is hardly compatible with continuous SU recording. From a technical point of view, we extracted as ‘MUA’ cells that were not reliably identifiable as SU but with clearly-defined action potentials, and only kept as occasional ‘SU’ cells that were readily separable according to their amplitude.
Electrically-evoked activity within LC and TCCWe adapted a previously described method to extract LC electrically-evoked activity [7]: post-stimulus time histogram (PSTH) were constructed using spikes evoked from desired stimulations, and bins of evoked activity were defined as bins with a frequency of discharge greater than the mean frequency of bins in the last 500 ms of the PSTH. The different components of the response were then identified and quantified as mean frequency of discharge above this last 500 ms epoch, and normalized by baseline frequency (i.e. divided by the mean frequency before the electrical train). A and C responses within TCC were extracted under 30 ms post-stimulus for an A response, and between 30–200 ms post-stimulus for a C response (PSTH were used to confirm in each case that these latencies were appropriate).
Mean frequency analysis (spontaneous activity)Mean frequency was computed using a centred window of defined width (depending on the analysis) at each spike event, and oversampled/decimated to a fix sampling rate. Some results are presented as a total activity evoked by a stimulation: for the TCC, it corresponds to the raw sum of the number of spikes during the appropriate time window, for the LC, it is the raw count of MUA divided by pre-stimulus count so that the result is a percentage of baseline activity.
Phase analysisMean MUA frequency was first bandpassed (FIR filter), and Hilbert transform was performed to extract the corresponding phase. Phase histograms were then constructed by allocating each spike to the corresponding bin of the phase during which it occurred (75 or 150 bins total for a period). Histograms were z-scored to keep only temporal dynamics and process mean over similar conditions.
Power in MUAThe ratio of power in the delta band (0.3 to 2 Hz) to total power of MUA was extracted on a mean frequency of MUA. To compare propranolol effect as before/after, a mean ratio was computed on 10 min between trains of electrical stimulations (two inter-train windows) either before dural injection and microinjection or after microinjection.
Local field Potential (LFP)Raw LFP signals were decimated (low pass filtered and down sampled) down to a sampling frequency of 50 Hz. Spectrograms of decimated signals were generated using Short Time Fourier Transform with the following parameters: Hamming window of length 1024, overlap of 1014 and FFT length of 8096. The log of resulting power spectral density (PSD) was then integrated from 0.3 Hz to 2 Hz to produce a time-varying value reflecting the low frequency activity within the signal. Correlations between this resulting power and the slow variations of MUA mean frequency (slow oscillations) were computed on 400 s sliding windows after smoothing (Savitsky-Golay, order 3, 51 samples), and R2 max was extracted for each rat.
Cross-correlograms were used to determine the mean lag between Sp5C and LC LFP signals, the peak giving the value and the direction of the lag.
StatisticsComparisons among groups for the behavioral studies were performed by 2-way analysis of variance (ANOVA). When only two sets were compared, normality was checked using Lilliefors test. If the hypothesis of normality was true, Student’s t test (paired or unpaired, according to conditions), otherwise Wilcoxon signed-rank test or Mann–Whitney U test were used.
Code availabilityComputer code used to analyze the datasets is available from the corresponding authors on reasonable request.
Data availabilityThe datasets analysed during the current study are available from the corresponding authors on reasonable request.
Comments (0)