
Remember me
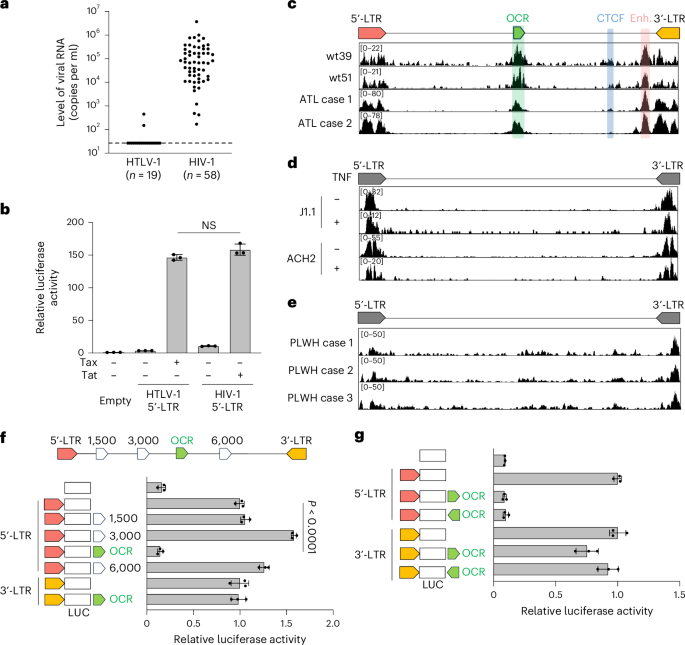

We compared plasma viral RNA levels in individuals infected with HIV-1 or HTLV-1. HIV-1-infected individuals showed significantly higher viral RNA levels than HTLV-1-infected individuals, whose viraemia was mostly undetectable (Fig. 1a and Supplementary Table 1). Only 2 of 19 HTLV-1 asymptomatic carriers had detectable viraemia, with no correlation to proviral load (PVL) (Extended Data Fig. 1a and Supplementary Table 2). Among various viral proteins, HIV-1 Tat and HTLV-1 Tax regulate proviral gene expression. Promoter assay using Jurkat T cells showed no significant difference in 5′-LTR transcriptional activity between HIV-1 and HTLV-1, regardless of Tat or Tax presence (Fig. 1b). This suggests that factors beyond these promoters, such as some other cis- or trans-elements, contribute to the stark contrast in virus productivity.
Fig. 1: Identification of an OCR in the middle of the HTLV-1 proviral genome with a suppressive function on the promoter activity of the HTLV-1 5′-LTR.
a, Plasma viral RNA levels in HTLV-1- or HIV-1-infected individuals. Viral RNA copy numbers were evaluated by reverse-transcription quantitative PCR (RT-qPCR) and ddPCR for HTLV-1 and HIV-1, respectively. b, The transcriptional activity of HTLV-1 or HIV-1 5′-LTRs was assessed by luciferase (LUC) reporter assay in Jurkat T cells with their respective trans-activators (Tax and Tat). c–e, ATAC-seq signals in the provirus region (5′-LTR, OCR, CTCF, enhancer (Enh.) and 3′-LTR) of HTLV-1-infected Jurkat T cell clones (wt39 and wt51) and PBMCs from two patients with ATL (c), HIV-1-infected T cell lines (J1.1 and ACH2) with or without TNF stimulation (d) and PBMCs from three HIV-1-infected individuals (people living with HIV-1, PLWH) (e). f,g, Effect of the OCR or three randomly selected proviral regions on the 5′-LTR or the 3′-LTR promoter activity. Jurkat T cells were used for luciferase reporter assay 48 h after transfection (f). The directionality of the OCR did not change the effect on the 5′-LTR or the 3′-LTR promoter activity (g). At least two independent experiments were performed. The bars and error bars represent the mean ± s.d. of results in triplicate experiments. P values were calculated using a two-sided, unpaired Student’s t-test (NS, not significant).
We hypothesized that an unidentified 5′ regulatory region in the HTLV-1 provirus suppresses the 5′-LTR promoter activity. To test this hypothesis, we performed an assay for transposase-accessible chromatin with sequencing (ATAC-seq) analysis to identify open chromatin regions (OCRs) in the provirus with HTLV-1-infected cells. We used JET cells, a Jurkat T cell line containing the tdTomato fluorescent reporter protein under the control of a Tax-responsive element20. These cells were infected with wild-type HTLV-1, and then cloning was performed by limiting dilution to obtain infected clones in a previous study17. These infected JET cell clones contained one copy of intact HTLV-1 provirus while sustaining the latent state (Extended Data Fig. 1b–e). We also analysed fresh peripheral blood mononuclear cells (PBMCs) from two patients with smouldering ATL, a clinical subtype of ATL with an indolent course. ATAC-seq identified two open chromatin peaks in the 3′ proviral region, corresponding to known insulator and enhancer elements17,18 (Fig. 1c). In addition, we discovered a previously unreported OCR in the 5′ proviral side, overlapping the viral polymerase coding region. Unlike HTLV-1, HIV-1 showed no intragenic ATAC-seq peaks beyond the LTRs, including in fresh CD4+ T cells from HIV-1-infected individuals (Fig. 1d,e). No similar intragenic OCRs were detected in other delta retroviruses, such as HTLV-2, simian T-lymphotropic virus 1 (STLV-1) or bovine leukaemia virus (BLV) (Extended Data Fig. 1f), although we cannot exclude the possibility of underestimation by using infected cell lines, in which the provirus tends to be silenced by DNA hypermethylation in the 5′-LTR promoter21. To investigate the function of the OCR in the HTLV-1 provirus, we performed a reporter assay using the 5′-LTR and 3′-LTR as promoters. The OCR significantly suppressed 5′-LTR promoter activity whereas other intragenic proviral regions did not (Fig. 1f). By contrast, the OCR showed little suppressive effect on the 3′-LTR, the promoter of antisense transcription from the HTLV-1 provirus (Fig. 1g). As the 5′-LTR is a TATA-box-containing promoter, whereas the 3′-LTR lacks an antisense TATA box, they show distinct promoter characteristics22,23. Consequently, the effect of the OCR should differ between them.
Molecular characterization of the silencer complex on the OCRTranscription factor (TF) motif analysis identified several candidate sites within the OCR, showing high similarity to consensus sequences for TF binding (Supplementary Table 3). Integrating ATAC-seq data, we selected RUNX, ETS and GATA TF families as key candidates for silencer function. Among them, RUNX1, ETS1 and GATA3 were most highly expressed in CD4+ T cells (Extended Data Fig. 2a–c). RUNX1 showed the strongest suppressive function in reporter assays (Extended Data Fig. 2d). Although other family members may contribute, these data suggest that RUNX1, ETS1 and GATA3 play major roles, leading us to focus on them in this study.
To assess binding to the OCR, we performed chromatin immunoprecipitation with sequencing (ChIP-seq) using viral DNA-capture-seq24. RUNX1, ETS1, GATA3 and the RUNX co-factor CBFβ bound to the OCR in both HTLV-1-infected cell lines and fresh PBMCs (Fig. 2a,b). RUNX proteins act in promoters, enhancers, insulators and silencers, with tissue-specific functions dictated by expression patterns, post-transcriptional modifications and stoichiometry of RUNX-mediated transcriptional complex25,26. Further analysis regarding cofactors related to RUNX1 revealed that HDAC3 and Sin3A localized to the OCR, strongly in PBMCs and marginally in cell lines (Fig. 2a,b), suggesting that they mediate OCR silencing via the RUNX1–CBFβ complex in naturally infected cells. HTLV-1-infected cell lines tend to show hypermethylation in the 5′-LTR, making them less dependent on silencer function to maintain a latent state21. Additional studies are needed to fully elucidate the molecular mechanisms underlying the silencer complex.
Fig. 2: Molecular characterization of the silencer complex on the OCR.
a,b, ChIP-seq signals with HTLV-1 DNA-capture analysis for RUNX1, CBFβ, GATA3, ETS1, HDAC3 and Sin3A in HTLV-1-infected Jurkat T cell clones (wt39 and wt51) (a) and PBMCs of patients with ATL (Al-5 and AI-9) (b). c, Effect of overexpression of RUNX1, GATA3 or ETS1 on the 5′-LTR promoter activity with the OCR in 293T cells. d, Effect of overexpression of RUNX1 and/or ETS1 on the 5′-LTR promoter activity with the OCR in 293T cells. e, Changes in OCR-mediated silencing by mutating RUNX, GATA3 and ETS1 binding sites within the OCR in Jurkat T cells. f, Transcriptional regulation of RUNX1 mutants (S67I, W79C and R174Q) in OCR-mediated silencing of HTLV-1 5′-LTR in 293T cells. Protein expression of RUNX1 mutants was confirmed twice by western blot. g, Representative RT-qPCR result of tax mRNA expression in MT1 and TBX-4B cells transduced with RUNX1. h, Measurement of Tax protein levels in RUNX1-overexpressed MT1 cells. RUNX1 was transduced using a retroviral vector system. One representative result from each flow cytometry assay (left) and the cumulative Tax positivity values from a triplicate assay (right) are shown. i, Effect of RUNX1 knockdown via shRNA on OCR-mediated silencing of the HTLV-1 5′-LTR. Molt4 cells carrying shRNA for RUNX1 were used for luciferase assay. Luciferase reporter assays were performed 48 h after transfection. The results are representative of at least two independent experiments. The bars and error bars represent the mean ± s.d. of the results of triplicate experiments. P values were calculated using a two-sided, unpaired Student’s t-test.
The OCR contains three RUNX1, ETS1 and GATA3 binding sites (Extended Data Fig. 3a). A core silencer region encompassing these sites showed silencing activity comparable to the full OCR (Extended Data Fig. 3b). We analysed TF effects on OCR-mediated suppression using a 5′-LTR reporter assay in 293T cells, which have lower endogenous TF expression than Jurkat cells. RUNX1 suppressed transcription, ETS1 enhanced it and GATA3 had little effect (Fig. 2c). These TFs also localized to the LTRs, but ChIP-seq could not distinguish between 5′- and 3′-LTR binding owing to their identical sequences (Fig. 2a,b). RUNX1 and ETS1 affected 5′-LTR promoter activity (Extended Data Fig. 3c,d), but their effects were stronger when the OCR was present, suggesting that their interplay modulates transcription. ETS1 counteracted RUNX1-mediated suppression (Fig. 2d). Mutations in all three RUNX1 sites abolished silencing, while ETS1 and GATA3 site mutations had no effect (Fig. 2e), highlighting the importance of RUNX1 as a key DNA-binding molecule to the OCR. Partial silencing loss occurred when individual RUNX1 sites were mutated, indicating that all three contribute (Extended Data Fig. 3e). Impaired silencer function from RUNX1 mutations could lead to aberrant viral protein expression and influence clinical outcomes, such as early ATL or HAM and tropical spastic paraparesis27. However, these RUNX1 sites were highly conserved across geographic strains (Extended Data Fig. 3f,g). Some ATL cases had OCR deletions, but these also lacked the 5′-LTR, suggesting that proviral silencing is maintained regardless of the OCR (Extended Data Fig. 3h,i).
HTLV-1 infects various cell types28 but primarily transforms CD4+ T cells. We hypothesized that the silencer function of the OCR is most active in CD4+ T cells, enabling viral persistence by evading immune detection. A reporter assay in different cell lines confirmed the strongest OCR-mediated suppression in T cells, not B cells or other lineages (Extended Data Fig. 4a). We additionally analysed the silencer function of the OCR on the 5′-LTR using primary CD4+ T cells and immortalized macrophage-like cells derived from induced pluripotent stem cells (Extended Data Fig. 4b).
To identify the RUNX1 domain responsible for silencing, we performed a 5′-LTR reporter assay with wild-type and mutant RUNX1 proteins29. Mutations in the DNA-binding Runt domain, disrupting DNA binding, transcriptional activity or CBFβ interaction, impaired silencing (Fig. 2f). Overexpressing RUNX1 in ATL and HTLV-1-infected cell lines reduced endogenous tax expression at both RNA and protein levels (Fig. 2g,h). Conversely, RUNX1 knockdown diminished 5′-LTR suppression in Jurkat and Molt4 cells (Fig. 2i and Extended Data Fig. 4c–e). Previously reported ATL-associated RUNX1 mutants retained silencer function (Extended Data Fig. 4f).
Virological and immunological significance of the OCRWe investigated the functional role of the OCR from a virological perspective. First, we identified mutations in the OCR that retained the pol gene coding sequence but lost transcriptional repression. Four nucleotide substitutions in RUNX1 binding sites abrogated silencing in the 5′-LTR promoter assay (Fig. 3a). We then generated a recombinant HTLV-1 with these mutations and established JET cells infected with either wild-type (wt) or silencer mutant (s-mut) HTLV-1 (Fig. 3b). Binding of RUNX1, CBFβ and HDAC3 to the silencer region significantly decreased in s-mut-infected cells (Fig. 3c). Consistently, tax expression was markedly upregulated in s-mut-HTLV-1-infected cells, whereas HBZ levels increased but less prominently (Fig. 3d).
Fig. 3: Virological and immunological significance of the OCR function in HTLV-1 infection.
a, The nucleotide and protein sequences of silent mutations (s-mut) in the RUNX binding site (underlined) within the OCR. Mutated nucleotides are shown in red. Luciferase reporter assays were performed using Jurkat T cells 48 h after transfection. b, Experimental workflow illustrating the establishment of reporter cells (JET cells) infected with wt- or s-mut-HTLV-1. c, Signals from RUNX1, CBFβ or HDAC3 ChIP-seq in the HTLV-1 provirus for wt- and s-mut-HTLV-1-infected JET cells. d, Representative RT-qPCR results showing tax or hbz mRNA expression levels in JET cells infected with wt- or s-mut-HTLV-1. e, Supernatant p19 ELISA results from JET cells infected with wt- or s-mut-HTLV-1 with or without PMA–ionomycin stimulation. f, PVLs during long-term culture in wt- or s-mut-HTLV-1-infected JET cells after tdTomato sorting. The data presented are representative of two independent experiments. g, Tax expression after treatment with RUNX1 inhibitor (Ro5-3335). Tax expression was analysed by expression of the reporter protein tdTomato. h, Effect of RUNX1 inhibitor on ex vivo Tax expression using PBMCs of ACs and patients with ATL. Tax-expressing CD4+ T cells were measured by flow cytometry. i, IFNγ ELISPOT assay results from Tax TCR-transduced CD8+ T cells cocultured with wt- or s-mut-HTLV-1-infected JET cells expressing HLA-A*24:02 (A24). At least two independent experiments were performed. The bars and error bars represent the mean ± s.d. of results in triplicate experiments. P values were calculated using a two-sided, unpaired Student’s t-test (n.d., not detectable).
Next, we analysed viral productivity. The s-mut virus produced significantly more virus in culture supernatants, while wt-HTLV-1 did not, even with phorbol 12-myristate 13-acetate (PMA)–ionomycin stimulation (Fig. 3e). To assess viral persistence, we cultured infected Jurkat T cells (Fig. 3b) for an extended period. The PVL decreased in wt-HTLV-1-infected cells but increased in s-mut-infected cells, consistent with higher viral gene expression and productivity while maintaining provirus as the dominant form of viral DNA (Fig. 3f and Extended Data Fig. 5a). In line with this, active histone modifications (H3K4me3, H3K9ac) at the 5′ side of provirus were reduced in wt- but not s-mut-HTLV-1-infected cells (Extended Data Fig. 5b). Chromatin conformation capture showed closer 5′-LTR-OCR proximity in wt- compared with s-mut-infected cells (Extended Data Fig. 5c,d), suggesting that the silencer complex influences epigenetic regulation of HTLV-1 in addition to the proviral insulator region.
Given the role of OCR in silencing, we examined whether it facilitates immune evasion by minimizing viral antigen expression. Using Ro5-3335, a RUNX1 inhibitor30, we assessed its impact on viral antigen expression. The drug showed no cytotoxicity at the tested concentration in HTLV-1- or HIV-1-infected cell lines (Extended Data Fig. 6a). In JET cell clones, Ro5-3335 significantly increased tdTomato expression in a dose-dependent manner (Fig. 3g and Extended Data Fig. 6b). As reported previously31, high-dose Ro5-3335 modestly increased HIV-1 proviral expression (Extended Data Fig. 6b,c). Tax expression, typically undetectable in freshly isolated PBMCs from HTLV-1-infected individuals but inducible by ex vivo culture32, was enhanced by Ro5-3335 (Fig. 3h and Extended Data Fig. 6d).
To determine whether OCR impairment increases susceptibility to cytotoxic T lymphocyte (CTL) responses, we compared Jurkat T cells infected with wt- or s-mut-HTLV-1. We used HTLV-1 Tax as the viral antigen for presentation by HLA-class I (HLA-A*24:02), a well-characterized and dominant HTLV-1 epitope33. We transduced the Tax-specific TCRs identified in the previous study34 into primary T cells and co-cultured with JET cells infected with the wt or the s-mut virus. We performed an enzyme-linked immunospot (ELISPOT) assay and found that infected cells with the s-mut virus showed higher immunogenicity than those with the wt virus (Fig. 3i and Extended Data Fig. 6e). Notably, the RUNX1 inhibitor treatment increased the susceptibility of infected cells against anti-viral CTL in both cytokine production and cytotoxicity assay by using anti-Tax-specific CTLs (Extended Data Fig. 6f,g). Although we cannot exclude the possibility that off-target effects of the RUNX1 inhibitor may contribute to the experimental outcome, these results establish the immunological significance of the OCR in controlling the viral antigen Tax.
Single-cell analysis of CD4+ T cells from people with HTLV-1Although HTLV-1-infected cells typically remain latent, the silenced provirus can be reactivated, triggering a transcriptional burst14,32 (Fig. 3h). This reactivation facilitates viral spread and de novo infection, although in vivo evidence is lacking. We cultivated primary CD4+ T cells from three indolent types of ATL case ex vivo overnight to induce transcriptional burst and performed multiome, simultaneous single-cell RNA sequencing (sc-RNA-seq) and sc-ATAC-seq analysis using cells both before and after cultivation. These patients with ATL contained the oligoclonal expansion of infected cells and retained the reactivation ability of tax expression after ex vivo culture (Extended Data Fig. 7a–d). Subsequently, we conducted cell clustering analysis by integrating sc-RNA-seq and sc-ATAC-seq data (Fig. 4a,b and Extended Data Figs. 8a,b and 9a,b). Infected cells were identified by their transcriptional characteristics (Fig. 4c and Extended Data Figs. 8c and 9c; see Methods for more details), and we further refined cell clustering using only the infected cells. Two distinct clusters emerged: one exclusively post-cultivation and another present both pre- and post-cultivation. The former corresponded to a transcriptional burst cluster, while the latter remained latent. Both clusters highly expressed CADM1, a marker of HTLV-1-infected cells35,36 (Fig. 4f and Extended Data Figs. 8f and 9f).
Fig. 4: Single-cell multiome analysis of CD4+ T cells from a smouldering ATL case (a5).
a,b, WNN UMAP projection of all cells, both before and after cultivation identified using multimodal neighbours by weighting a combination of ATAC and RNA data. Cells are labelled by cell type (a) and before or after cultivation (b). c, Heatmaps of expression levels of viral RNA and CADM1, a marker of HTLV-1-infected cells. d,e, WNN UMAP projection of only HTLV-1-infected cells, both before and after cultivation identified using multimodal neighbours by weighting a combination of ATAC and RNA data. Cells are labelled by cell type (d) and before or after cultivation (e). f, Heatmaps of expression levels of viral RNA and CADM1, a marker of HTLV-1 infected cells. g, Aggregated and normalized ATAC signals of burst and latent cell clusters in the proviral region. h, Violin plots of viral RNA expression in burst and latent cell clusters. i, Violin plots of infected cells and UMAP projections of all cells with a heatmap showing the RNA expression level of transcription factors related to the OCR in burst and latent cell clusters. j, Heatmaps of RUNX1 motif activity and a correlation plot showing the relationship between the sense transcription of HTLV-1 (horizontal axis) and the motif activity of RUNX1 (vertical axis). The blue line represents the regression line. The plot is labelled for burst and latent clusters. k, Schematic figure of the OCR-mediated silencing in HTLV-1 provirus.
ATAC-seq analysis revealed high chromatin openness across the HTLV-1 proviral region in the burst cluster, consistent with active transcription (Fig. 4g–h, Extended Data Figs. 8g–h and 9g–h). By contrast, the latent cluster showed three ATAC-seq peaks between the 5′- and 3′-LTRs, aligning with known regulatory regions: the OCR, insulator18 and enhancer17.
We further examined gene expression dynamics of transcription factors during transcriptional bursts using sc-multiome data. Expression of RUNX1 and ETS1 decreased, whereas that of GATA3 and Sin3A increased (Fig. 4i and Extended Data Figs. 9i and 10i). Even though the expression level of RUNX1 was decreased, there was a moderate decrease in RUNX1 TF motif activity in the burst cell population (Fig. 4j and Extended Data Figs. 9j and 10j), indicating that changes in expression of cofactors, such as ETS1 and GATA3, can be key determinants for the silencer function. In particular, ETS1 strongly upregulates transcriptional activity mediated by the 5′-LTR and OCR, making it a crucial molecular switch for RUNX complex function (Fig. 2c)15. Further studies are needed to elucidate how intragenic regulatory elements—silencer, insulator and enhancer—coordinate proviral expression and retroviral latency in the host.
Effect of the HTLV-1 OCR on the virological aspect of HIV-1We tested the hypothesis that the RUNX1-mediated HTLV-1 silencer functions as a dominant silencer in both HIV-1 and HTLV-1 (Fig. 1a). Here we aimed to analyse the effect of the OCR on the HIV-1 5′-LTR (Fig. 5a) by constructing a novel recombinant HIV-1 (rHIV-1) molecular plasmid clone. This involved inserting either the wt or the mutated OCR into the HIV-1 nef region (Fig. 5a,b), which is a well-established region for the insertion of external genes of interest, such as reporter fluorescent protein37. Although this region is distant from the 5′-LTR in the HIV provirus, chromatin looping has been reported between the 5′- and 3′-LTRs during active transcription of HIV-1 (ref. 38), suggesting that the OCR in the nef region may suppress HIV-1 5′-LTR activity.
Fig. 5: Introduction of HTLV-1 OCR into rHIV-1 decreases proviral expression, virus production and cytopathic effect.
a, Schematic representation of the rHIV-1 construction with wt- or mt-OCR derived from HTLV-1. The NL4-3 plasmid was used to generate the rHIV-1 constructs. b, Effect of HTLV-1 wt- or mt-OCR on the HIV-1 5′-LTR promoter activity. Luciferase reporter assays were performed using Jurkat T cells 48 h after transfection. c, Quantification of cell-associated viral DNAs after infection with each rHIV-1. gDNAs were extracted from Jurkat T cells infected with each rHIV-1. d, RUNX1 ChIP–qPCR analysis of Jurkat T cells infected with rHIV-1 with wt- or mt-OCR. Enhancer RUNX1 region (eRUNX1) was used as a positive control for ChIP. e, HIV-1 production in Jurkat T cells infected with rHIV-1. Intracellular HIV-1 p24+ cells were detected using flow cytometry. f, Assessment of the cytopathic effect induced by rHIV-1. Cell viability and dead cell counts were determined using the trypan blue exclusion method. g, A representative workflow and flow cytometry results of the WIPE assay using each rHIV-1 are presented. At least two independent experiments were performed. The bars and error bars represent the mean ± s.d. of results in triplicate experiments. P values were calculated using a two-sided unpaired Student’s t-test.
Using the NL4-3 plasmid39, we generated rHIV-1 containing either wt- or HTLV-1 silencer mutant (mt)-OCR and conducted single-round infections in Jurkat T cells. One day postinfection, intracellular HIV-DNA levels showed no significant differences among the viruses (Fig. 5c). RUNX1 ChIP-qPCR confirmed RUNX1 binding to the inserted silencer region in the integrated provirus (Fig. 5d). The level of intracellular p24 gag expression at 2 days postinfection was dramatically decreased in the rHIV-1 with the wt-OCR but not that with the mt-OCR lacking RUNX1 binding capacity (Fig. 5e). Notably, the cytopathic effect induced by viral infection was significantly decreased by insertion of the wt-OCR but not by the mutated OCR (Fig. 5f).
To assess the role of OCR in viral persistence, we used the widely distributed intact provirus elimination (WIPE) assay40. Introduction of the OCR remarkably decreased viral persistence in the WIPE assay, but the effect was partially cancelled by mutation in the RUNX-binding sites (Fig. 5g). These findings show that OCR exerts a dominant silencing effect on HIV-1, highlighting its role in shaping distinct infection outcomes for HIV-1 and HTLV-1.
Comments (0)