Several studies reported increased uptake of FAP ligands in infarcted hearts post -MI, and correlation with clinical findings and magnetic resonance imaging (MRI) markers of myocardial damage, and even prediction of late LV remodeling [12, 13, 25]. Notably, FAP-expression was not only detected within the infarct scar, but also within the surviving myocardium as indicated by lack of direct alignment with late gadolinium enhancement (LGE) territory on MRI or the perfusion defect defined by SPECT [12, 13, 25]. This corresponds to histological findings in rat hearts demonstrating FAP expression extends beyond the infarct scar to the surviving myocardium near the necrotic area post-MI [10, 16]. To reconcile these findings, we characterized FAP-expressing cell distribution in necrotic and surviving myocardium post-reperfusion of experimental MI/R.
Identification of the AAR after MI/R
We defined the AAR by absence of endothelial staining using in vivo lectin infusion to label capillaries and large vessels with high resolution [21]. Similar methods have been described including fluorescent microsphere injection and 2,3,5-triphenyltetrazolium (TTC) staining [6, 26]. Non-invasive radionuclide imaging of AAR faces spatial resolution limitations, which can be addressed in small animal models by ex vivo autoradiography. Differences in infarct and AAR have been characterized by multi-tracer autoradiography in a reperfused infarct rat model using thallium-201 and 99mTc-Sestamibi radiotracers. Consistent with our study, autoradiography defined a remarkably larger AAR compared to the infarct scar [27].
In humans, AAR can be estimated non-invasively by cardiac MRI, PET imaging or (99 m)Technetium (Tc)-Sestamibi SPECT [28,29,30]. However, these non-invasive methods are not performed routinely before coronary reperfusion in STEMI, making it challenging to identify initially perfused vs non-perfused myocardium retrospectively. In our study, in vivo lectin staining enabled precise AAR delineation post-MI/R using multi-color immunofluorescence and high -resolution microscopy surpassing the spatial resolution of current non-invasive techniques in humans.
Areas of hypoxia were indeed confined to the AAR, but the AAR was not exclusively hypoxic, especially at 3 days post-MI/R. Concordantly, hypoxic area was smaller than ischemic myocardium in individual animals. One explanation is that pimonidazole detects hypoxia at pO2 below 10 mmHg, whereas normal capillary bed pO2 is as low as 40 mmHg [31, 32]. An oxygen gradient within the infarct borderzone likely cannot detect intermediate hypoxia (10–40 mmHg) [33, 34], consistent with the narrow non-hypoxic transition zone at the border of ischemic and non-ischemic myocardium in our study. Moreover, early neovascularization of the reperfused myocardium might have improved oxygen supply to the scar despite LAD-occlusion [35].
However, we clearly identified the AAR by visualizing ischemic and non-ischemic myocardium. We further showed that hypoxic area is smaller than AAR, which can be reconciled given the known spatial and temporal heterogeneity of tissue oxygen in pathological conditions and neovascularization [32, 34].
FAP Expression and Inflammatory Cell Infiltration Increase within the AAR After MI/R
Our study shows that FAP+ myofibroblasts and CD68+ macrophages infiltrate the infarcted heart with a peak at 3 days post-MI/R, often in close proximity but as distinct populations. This relationship may stem from myofibroblast activation via cytokines such as interleukin-1, −6, and TNF-α released by recruited macrophages and monocytes [3, 36]. MI/R leads to inflammatory cell invasion and myofibroblast activation, and requires tight temporospatial cell coordination for effective scar maturation [37, 38]. Ultimately, the healing infarct consists of cell debris and granulation tissue containing mostly inflammatory cells, myofibroblasts and endothelial cells [2, 38]. Supporting our findings, FAP+ fibroblasts and macrophages were also upregulated and co-localized in colorectal cancer tissues, where a synergistic cell–cell interaction network between these cell populations was identified [39].
FAP expression was non-uniformly distributed within the AAR, confined to areas with activated myofibroblasts in the scar and borderzone. In this respect, van den Borne et al. showed myofibroblasts occupy only ~ 5–10% of the infarct scar in mice [40]. Our results are also consistent with FAP expression in the infarcted area and borderzone after MI in rats [10, 16]. FAP+ cells were rarely detected in remote myocardium in both studies [10, 16].
Together, we show that FAP-expressing myofibroblasts are non-uniformly distributed within the AAR, with large FAP-negative regions.
FAP Expression Extends Beyond the Infarct Scar After MI/R
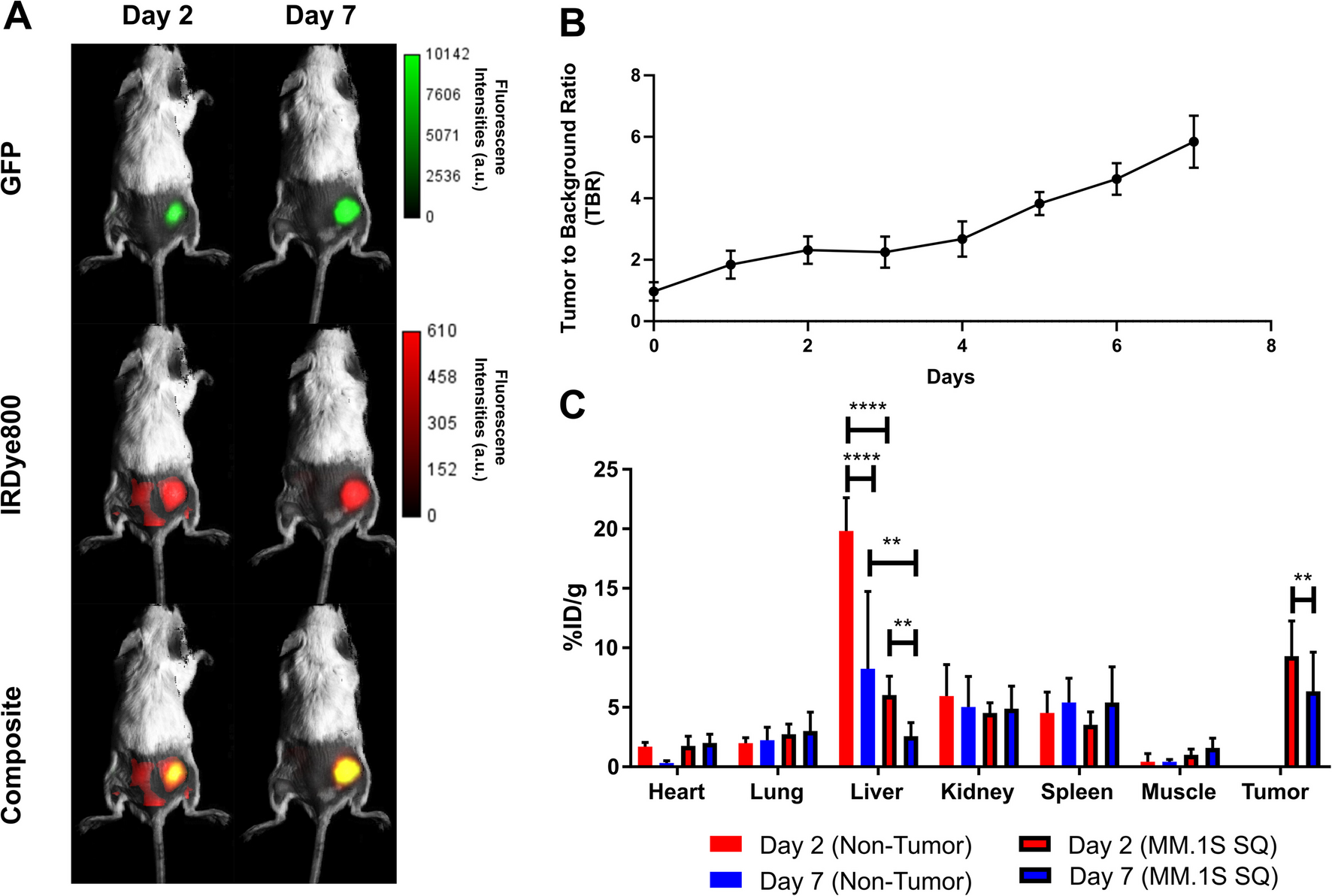
Immunohistochemistry showed FAP+ myofibroblast area was smaller than the AAR or hypoxic myocardium, with no correlation between these areas and FAP expression. This reflects that FAP was expressed only in activated myofibroblasts, while other cells like inflammatory cells and cell debris remained FAP negative. Moreover, FAP expression was evident in the scar, in surviving ischemic myocardium and sometimes extended into surviving myocardium adjacent to the infarct borderzone, as evidenced by autoradiography. However, in contrast to immunohistochemical analysis the area of myocardium with 68 Ga-FAPI-46 uptake was larger than the scar area after MI/R. A similar uptake pattern was previously reported with 68 Ga-FAPI-04 [16].
Despite similar non-uniform distribution, extent of FAP expression detected by immunohistochemistry and 68 Ga-FAPI-46-PET imaging remains discrepant. This might be explained by several factors: First, the lower spatial resolution of PET imaging compared to immunohistochemistry may contribute to the apparent larger radiotracer uptake area, especially given that the infarcted ventricle wall thickness in mice tends toward the resolution limits. This issue is compounded by the high positron range of gallium-68 which may complicate definition of the boundary of the infarct and border zone. Notably, these complexities are mitigated in part in clinical application with larger structures. Second, the 68 Ga-FAPI-46 probe may be more sensitive to extracellular FAP, detecting soluble FAP in low concentrations that immunohistochemistry may miss, resulting in a broader detection range by PET imaging especially near infarct margins [41]. These differences highlight the need for further studies to clarify the mechanisms underlying the varying detection of FAP across these methodologies.
Increased FAP expression within the AAR, borderzone and adjacent non-ischemic surviving myocardium can be explained by cytokine release post-MI/R. The borderzone, adjacent to the infarct, involves intact tissues with ischemia-specific transcriptomic changes [42]. This area is clinically relevant, since it can extend into non-injured myocardium during post-MI remodeling [4, 42, 43]. Diekmann et al. found FAP uptake correlated with perfusion defect size by SPECT and LGE by MRI, but the area of FAP uptake was about twice as large, indicating that 68 Ga-FAPI-46 uptake provides a measurement distinct from perfusion defect or tissue fibrosis. Moreover, FAP expression and LGE signals often mismatched in segmental infarct analysis, supporting a distinction between FAP+ cells and tissue fibrosis [13].
Thus, FAP may serve as a potential marker for myocardial injury extent and healing, complementing MRI, PET, and SPECT in humans, particularly targeting transient activated myofibroblasts in the early stage of disease [13, 16, 25, 44].
FAP is Upregulated After Acute MI in Humans in Perivascular and Reparative Fibrotic Tissue
Lastly, we demonstrate that FAP+ fibroblasts are present at low frequency within surviving areas near the infarct scar in humans with acute MI, consistent with previous findings [22]. The expression of CD13 demonstrated the spatial localization of FAP expressing cells in relation to fibroblast activation, inflammatory response and angiogenesis within the infarct and border zones during myocardial healing post-MI/R.
Recent studies highlighted the therapeutic potential of FAP + myofibroblasts, wherein cardiac fibrosis after angiotensin II and phenylephrine infusion was nearly eliminated by ablating FAP+ cells with antigen-specific CD8+ T cells [45].
Using clinical imaging we demonstrate that the myocardial area showing FAP upregulation exceeded the infarct region in patients shortly after acute MI and standard reperfusion therapy consistent with prior reports [12, 13]. The region of fibroblast activation on 68 Ga-FAPI PET extended beyond the perfusion defect visible with conventional SPECT. This finding was supported by cellular FAP expression outside the infarct area using immunohistochemistry in acute MI patients. This suggests that FAP upregulation plays a role in both replacement fibrosis in the injured area and reactive fibrosis in non-infarcted myocardium. A first report has shown a prognostic role of PET-derived fibroblast activation extent for later LV dysfunction [13].
Study Limitations
Our study was performed in mice, which exhibit variable wound healing responses post-MI depending on strain and gender [7, 40]. However, our results are consistent with findings in rats [10], and we showed that also in humans FAP+ cells were occasionally found outside the infarcted area in acute MI. We observed variability of ischemic areas between animals, and statistical significance was not reached likely due to due to the small sample size. Nevertheless, we demonstrated the unique pattern of FAP expression in ischemic and non-ischemic myocardium extending beyond the necrotic scar. Another limitation of our study is the lack of long-term timepoints for evaluation of FAP expression. Still, our study found maximum FAP expression at 3 days post-MI/R, and we previously demonstrated that FAP activity decreases afterwards [10]. Therefore, FAP distribution within the AAR would likely be smaller than observed at 3 days post -MI/R at later timepoints.




Comments (0)