
Remember me




Uveitis encompasses a spectrum of intraocular inflammatory conditions targeting that mainly the uvea, i.e., iris, ciliary body and choroid, as well as adjacent structures, including retina and vitreous [1]. It arises from an inappropriate activation of the immune system, leading to ocular inflammation that, if left unchecked, can result in irreversible damage to the sensitive components of the visual system [2]. These conditions represent significant global health burdens, contributing to visual impairment and blindness, particularly in developing nations. Understanding the mechanisms underlying ocular inflammation and its association with these ocular diseases is crucial for the development of effective therapeutic interventions.
Th17 cells, the IL-17 producing CD4+ T helper cells, are formed by heterogeneous subsets with different regulations and functions and have gained significant attention due to their crucial role in immune-mediated diseases, including uveitis [2,3,4,5]. Th17 cells are characterized by the production of signature cytokines including interleukin (IL)−17, IL-17F, IL-21, and IL-22 and the expression of the master transcription factor, retinoic acid–related orphan receptor (ROR)- γt [6]. Under specific microenvironment, they secrete addition proinflammatory cytokines, such as granulocyte–macrophage colony-stimulating factor (GM-CSF) [7, 8] and interferon (IFN)-γ [9] for tissue damage. Originally identified as key players in host defense against extracellular pathogens, Th17 cells and their signature cytokines have since been implicated in the pathogenesis of various autoimmune and inflammatory disorders, including ocular autoimmune diseases [2, 10].
In the context of autoimmune uveitis, Th17 cells have emerged as important mediators of immune dysregulation [11, 12]. These cells are involved in the initiation and perpetuation of inflammatory responses within the eye as demonstrated in patients with uveitis as well as the animal model, experiment autoimmune uveitis (EAU), which is usually induced by two retinal antigen, namely interstitial retinol–binding protein (IRBP) and S-antigen [2, 13]. Th17 cells are recruited to the site of inflammation in the eye, where they secrete pro-inflammatory cytokines, including IL-17A, IL-17F, GM-CSF, and IL-22 [11, 14, 15]. These cytokines contribute to the recruitment and activation of other immune cells, such as neutrophils and macrophages, and promote the production of additional inflammatory mediators [2, 14]. The activity of Th17 cells and their cytokines can lead to tissue damage and disruption of the blood-retinal barrier, further exacerbating ocular inflammation. Targeting Th17 cell–associated pathways and cytokines may offer new therapeutic possibilities for modulating ocular inflammation and improving visual outcomes in these conditions. Further research into the intricate interplay between Th17 cells and ocular inflammation will undoubtedly shed light on potential therapeutic strategies for managing ocular diseases with an inflammatory component [2].
In this review, we aim to provide a comprehensive overview of Th17 response in the pathogenesis of uveitis. We will discuss the underlying mechanisms of its cytokine milieu in ocular inflammation. Furthermore, we will explore potential therapeutic targets and interventions aimed at modulating Th17 response to prevent or treat autoimmune uveitis. Contrary to its conventional classification as a promoter of inflammation, some studies have unveiled a more nuanced role for Th17 cells. They appear to have the ability to produce immunoregulatory cytokines, particularly IL-10 and IL-24 [5, 9], and IL-17 increases the number of regulatory T cells (Tregs) [16]. This suggests that Th17 response may have a bifunctional role that encompasses both immune activation and suppression. By elucidating the intricate relationship between Th17 response and autoimmune uveitis, we hope to contribute to the development of innovative approaches for managing ocular autoimmune diseases and improving visual outcomes.
Th17 Response and Its Role in InflammationTh17 response refers to the activation and expansion of CD4+ T helper cells that produce IL-17 and other pro-inflammatory cytokines. Th17 cells play a critical role in immune responses against pathogens and is involved in the pathogenesis of various inflammatory diseases. The primary function of the Th17 response is to provide protection against extracellular pathogens, particularly fungi and bacteria [17]. IL-17 and other cytokines produced by Th17 cells recruit and activate neutrophils, which are crucial for combating these types of infections [18]. Th17 cells also stimulate the production of antimicrobial peptides and chemokines, further enhancing the immune response at the site of infection. However, dysregulation of the Th17 response can contribute to the development of chronic inflammatory diseases. Excessive or sustained Th17 activation can lead to tissue damage and perpetuate inflammation. Aberrant Th17 responses have been implicated in autoimmune disorders such as rheumatoid arthritis, psoriasis, and inflammatory bowel disease [17]. In these conditions, Th17 cells infiltrate affected tissues and release pro-inflammatory cytokines, leading to immune cell recruitment, tissue destruction, and chronic inflammation. In addition to autoimmune diseases, the Th17 response has been implicated in other inflammatory conditions, including neuroinflammatory disorders, such as multiple sclerosis, and allergic diseases, such as asthma. Th17 cells and their cytokines can contribute to the breakdown of the blood–brain barrier in neuroinflammation and promote the recruitment of immune cells into the central nervous system [19]. In asthma, Th17 cells are involved in airway inflammation and the recruitment of eosinophils, contributing to bronchial hyperresponsiveness [20, 21]. Understanding the role of the Th17 response in inflammatory diseases is essential for developing targeted therapeutic strategies. Modulating Th17 cell activity or targeting Th17-associated cytokines may offer potential treatment options for managing these conditions [10]. However, it is important to note that the Th17 response is a complex and dynamic process, and further research is needed to fully unravel its intricacies and exploit the therapeutic potential of targeting Th17 response in inflammation.
Th17 Cells: The Driving Force Behind Uveitis PathogenesisHuman autoimmune uveitis has been associated with the presence of Th17 response as supported by the elevation of IL-17 serum level and the expression of IL-17 from peripheral blood mononuclear cell (PBMC) in patients with uveitis and more importantly, high IL-17 level is associated with active uveitis and disease severity [4, 22, 23]. In addition, a genome-wide study on Bechet’s disease has identified that regions encompassing IL-10 and IL23R-IL12RB2, which are highly involved in Th17 differentiation, are associated with disease susceptibility [24]. To further elucidate the role of Th17 cells in the pathogenesis of uveitis, researchers have turned to animal models. Experimental autoimmune uveitis (EAU) is commonly induced in these models by immunizing animals with a retinal protein and bacterial adjuvants, such as pertussis toxin and complete Freund’s adjuvant [2].
The pivotal discovery that IL-23, rather than IL-12, is essential for the development of autoimmune diseases in animal models [25,26,27], has shifted the focus towards the Th17 response. This shift underscores the importance of Th17 cells, rather than Th1 cells, as the principal drivers of EAU pathogenesis [4, 11]. Luger et. al. reported that targeting IL-17, the defining cytokine produced by Th17 cells, markedly reduced ocular inflammation in mice with EAU. Furthermore, their research demonstrated that merely transferring Th17 cells predisposed to induce uveitis is sufficient to penetrate the retinal barrier and trigger the onset of EAU [11]. A follow-up study underscored the pivotal role of Th17 cells in the development of EAU. The study revealed that conditional knocking out STAT3, a key transcription factor necessary for the differentiation of Th17 cells [28], in CD4+ cells led to a deficiency of IL-17-producing CD4+ T cells in mice immunized for EAU. Consequently, these mice did not develop the disease, highlighting the essential contribution of Th17 cells to the pathogenesis of EAU [29]. The activation of uveitogenic Th17 cells has been associated with overactivation of innate components, including macrophages, dendritic cells, neutrophils, γδT cells, and natural killer cells [30,31,32]. It has been well demonstrated that proinflammatory macrophages and dendritic cells expressed high level of IL-6, IL-23, and other co-stimulatory molecules to promote the pathogenic Th17 cells, while Th17 cells activate them reciprocally by the expression of GM-CSF [33]. In addition, the essential role of the gut microbiome has been highlighted in the induction and activation of retinal-IRBP-specific Th17 cells in the gut of R161H mice, which spontaneously develop EAU [34]. A subsequent study in patients with Behcet’s uveitis also confirmed that their gut microbiome has a higher potential to induce IL-17 production from their PBMCs [35]. These suggest the complexity of regulating Th17 response during the pathogenesis of uveitis.
Following the elucidation of Th17 cells’ involvement in EAU, research efforts have pivoted towards targeting the Th17 pathway to mitigate this condition. The differentiation of Th17 cells is contingent upon the presence of two pro-inflammatory cytokines: IL-6 and IL-23 [6]. Strategies that involve neutralizing these cytokines with antibodies or inhibiting their receptors have been demonstrated to alleviate EAU [11, 36, 37]. Later, IL-1β has also been identified as a critical factor in promoting the differentiation of pathogenic Th17 cells [38]. This pivotal role of IL-1β has similarly been observed in the context of EAU [39]. More cytokines have been reported to ameliorate EAU by limiting the differentiation of Th17 cells, e.g., IFN-γ, IFN-α2a, IFN-β, and IL-27 or inhibiting the effector functions of Th17 cells, e.g., IL-10, IL-21, IL-24, and IL-35 [5, 39,40,41,42,43,44,45]. These findings suggest that cytokine-based immunotherapy aimed at modulating the differentiation and activity of Th17 cells represent a potential avenue for the development of effective treatments for uveitis.
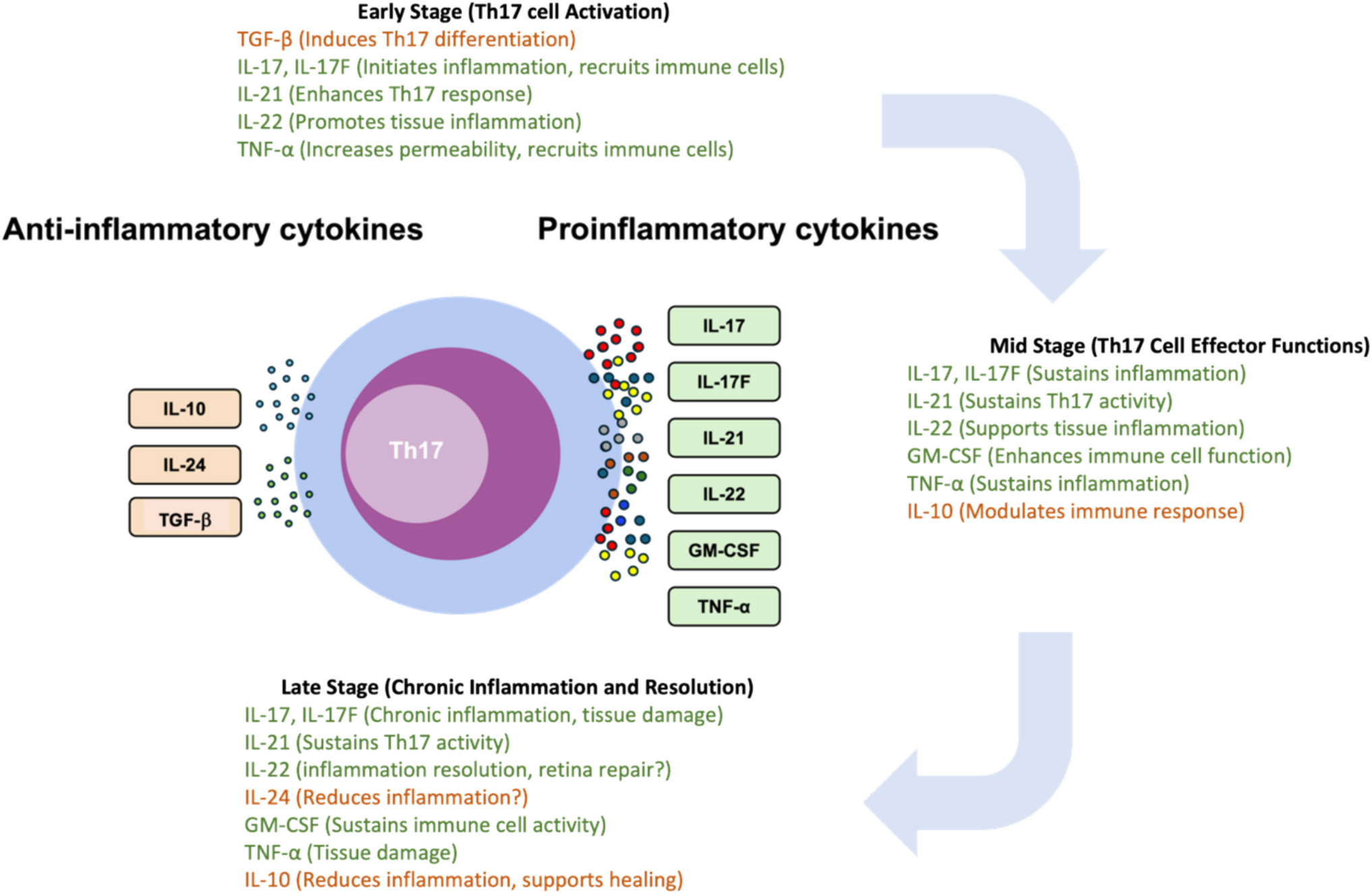
Th17 cells orchestrate inflammatory responses and tissue damage primarily through the secretion of proinflammatory cytokines, including IL-17, IL-17F, IL-21, IL-22, and GM-CSF [6] (Fig. 1). IL-17 and IL-17F are critical for mediating inflammation. Both IL-17 and IL-17F bind to the same IL-17RA/RC heterodimer receptor, although IL-17F has a much lower binding affinity and exhibits relatively weaker biological potencies compared to IL-17 [46]. Consequently, IL-17 is more prominently studied for its role in autoimmune diseases, including uveitis. They stimulate various cell types, including fibroblasts, endothelial cells, epithelial cells, and immune cells, to produce additional proinflammatory cytokines and chemokines for the recruitment of neutrophils and other immune cells to sites of inflammation [17]. In retina, IL-17 has been shown to promote proinflammatory cytokines production from retinal pigment epithelial (RPE) cells, astrocytes, and retinal microglia [17, 42, 47,48,49] and negatively affect retinal ganglion cells and other neuronal cells, potentially leading to vision impairment. In addition, IL-17 has implicated in the damage of blood-retinal barrier (BRB) by compromising the production of tight junction proteins from RPE cells and endothelial cells [19, 50].
Fig. 1
Cytokine profile of Th17 cells. Th17 cells are known to secrete pro-inflammatory cytokines that contribute to the pathogenesis of autoimmune conditions such as uveitis. Yet, these cells also have the capacity to produce anti-inflammatory cytokines, such as IL-10 and IL-24 and TGF-β, which have been associated with the resolution of immune responses
IL-21, as one of the Th17 cytokines, is implicated in the pathogenesis of a spectrum of proinflammatory and autoimmune disorders [51]. Elevated levels of IL-21 have been detected in the serum and aqueous humor of patients diagnosed with uveitis, suggesting a role in ocular autoimmune processes [52]. In EAU, CD4+ T cells that produce IL-21 have been observed infiltrating the eyes during the development of the disease. Remarkably, attenuation of IL-21 expression in these cells has been demonstrated to reduce their pathogenic potential, leading to a decrease in ocular inflammation. This finding underscores the importance of IL-21 in the exacerbation of inflammatory responses within the eye [53].
IL-22, belonging to the IL-10 cytokine family, is recognized for its pivotal role in tissue regeneration and the defense of barrier surfaces [54]. Its regulatory functions are essential for maintaining the integrity of tissue barriers. Consequently, IL-22 has been associated with various conditions characterized by inflammatory tissue pathology [55]. Its role in uveitis, however, presents a paradox. Although there is a study reported the elevation of serum IL-22 level in patients with acute anterior uveitis [56] along with findings of IL-22-producing CD4+ Th cells in ocular specimens from patients with Behçet’s disease [57], several studies suggest a lack of positive correlation between IL-22 levels and uveitis, reporting either no association or a negative one [58,59,60]. Despite these conflicting observations in clinical settings, EAU studies have shed light on a potentially protective role for IL-22. IL-22 treatment has been shown to resolve ocular inflammation by promoting the regulatory CD11b+ antigen presenting cells (APCs) [60]. This protective mechanism aligns with findings from another research, indicating that IL-22 may act locally within the retina to diminish inflammatory damage by enhancing the suppressive functions of retinal glial Müller cells [61]. The precise function of IL-22 in the context of autoimmune uveitis remains to be fully delineated. Additional research is essential to characterize the cytokine’s exact mechanisms of action and its potential dual roles in this complex ocular condition.
GM-CSF plays an essential role in the development, maturation, and activation of immune cells, especially myeloid cells including neutrophils and monocytes/macrophages [62]. Consequently, it is implicated in the onset and progression of inflammatory and autoimmune disorders [62]. The importance of GM-CSF in Th17 cell–mediated neuroinflammation has been highlighted in experimental autoimmune encephalomyelitis (EAE), the animal model of multiple sclerosis. It has been shown that Th17 cells can instigate EAE through the action of GM-CSF alone, even in the absence of the traditionally recognized pro-inflammatory cytokines, IL-17 and IFN-γ [7, 8]. Elevated level of GM-CSF has also been reported in the aqueous humor of patients with uveitis, suggesting its role in causing retina inflammation [52, 63]. In the case of EAU, So et. al. reported that IL-17 and IFN-γ double knockout mice are still capable to develop full prone retina inflammation with the ocular infiltrates contained increased GM-CSF-producing CD4+ T cells, in which GM-CSF propels the disease pathology by activating and drawing eosinophils into the ocular tissue [64].
Targeting Th17-related Cytokines in UveitisStrategies to mitigate Th17 cell–mediated inflammation have been developed, focusing on either inhibiting their differentiation and activation by depleting Th17-driving cytokines or neutralizing Th17 effector cytokines, as summarized in Table 1. The differentiation of Th17 cells from naïve CD4+ T cells is critically dependent on IL-23, IL-6, and IL-1β. Interventions aimed at these cytokines or their receptors have successfully suppressed Th17 cell proliferation and their cytokine production, thereby aiding the control of ocular inflammation in EAU [11, 12, 36, 37]. Ustekinumab, which blocks the Th1 and Th17 responses by targeting IL-12 and IL-23, has shown promise in treating spondyloarthritis-associated uveitis [65]. Additionally, Sarilumab, designed to inhibit Th17 cell differentiation by obstructing the IL-6 receptor, has been report to be effective in a phase 2 clinical trial for non-infectious uveitis [65, 66]. Anakinra, the IL-1 receptor antagonist, and canakinumab, the anti-IL-1β neutralization antibody, have shown improvement of retinal vasculitis lesions and uveitis flares with reduced steroid dosage in Behçet’s disease–related uveitis [67]. However, the efficacy of IL-17 neutralizing agents, such as ixekizumab and secukinumab, in treating uveitis has shown variable outcomes [10, 65, 68, 69]. Although ixekizumab display therapeutic effect on ankylosing spondylitis, it does not show any improvement in the related uveitis [70]. Three clinical trials of secukinumab in non-infectious uveitis did not meet the primary efficacy endpoints [71]. Collectively, these clinical studies suggest that while targeting Th17 cells is promising, depleting its signature cytokine, IL-17, yields discouraging outcomes. This indicates that IL-17 may have multiple roles in the pathogenesis of uveitis [10, 65, 68,69,70,71].
Table 1 Targeting Th17-related cytokines in uveitisAn Immunoregulatory Role of Th17 Cells in Attenuating Immune ResponsesThe involvement of the Th17 response in the pathogenesis of autoimmune diseases, primarily through the secretion of proinflammatory cytokines, is well-established. However, a growing body of evidence suggests a more nuanced role for Th17 cells beyond their proinflammatory functions. Notably, Th17 cells have been shown to express immunoregulatory cytokines, including IL-10, IL-24, and transforming growth factor (TGF)-β (Fig. 1) [5,
Comments (0)