
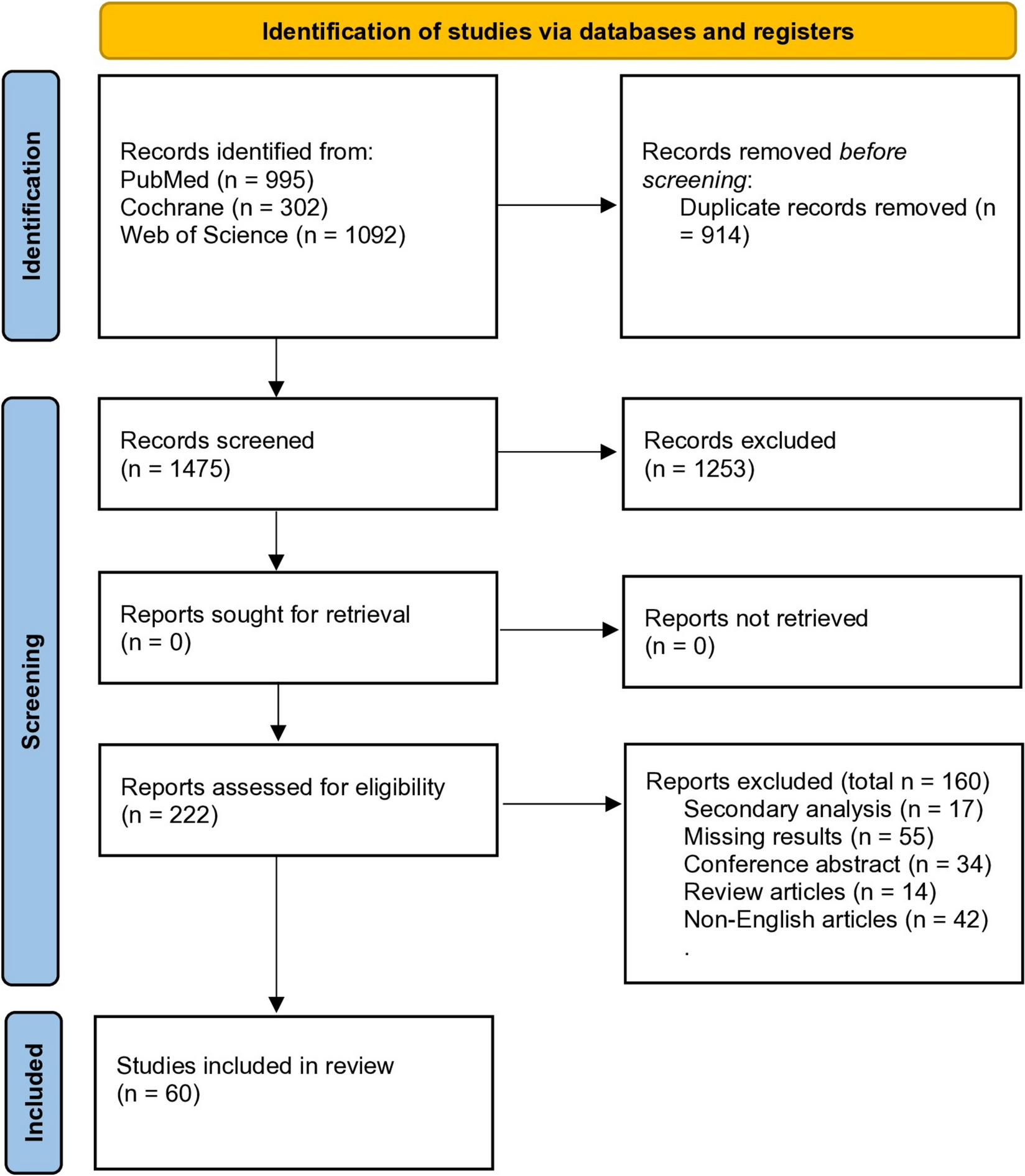
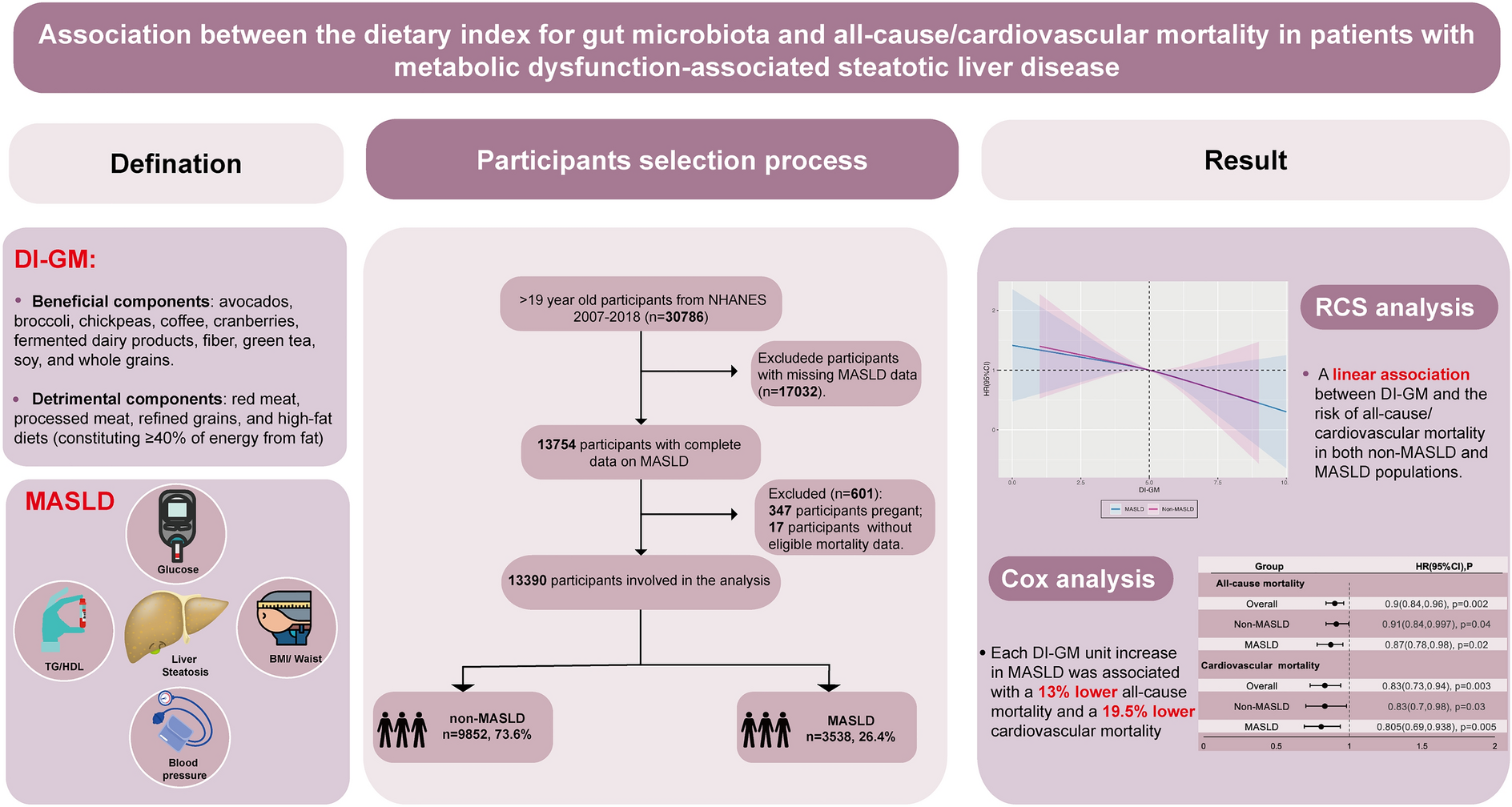
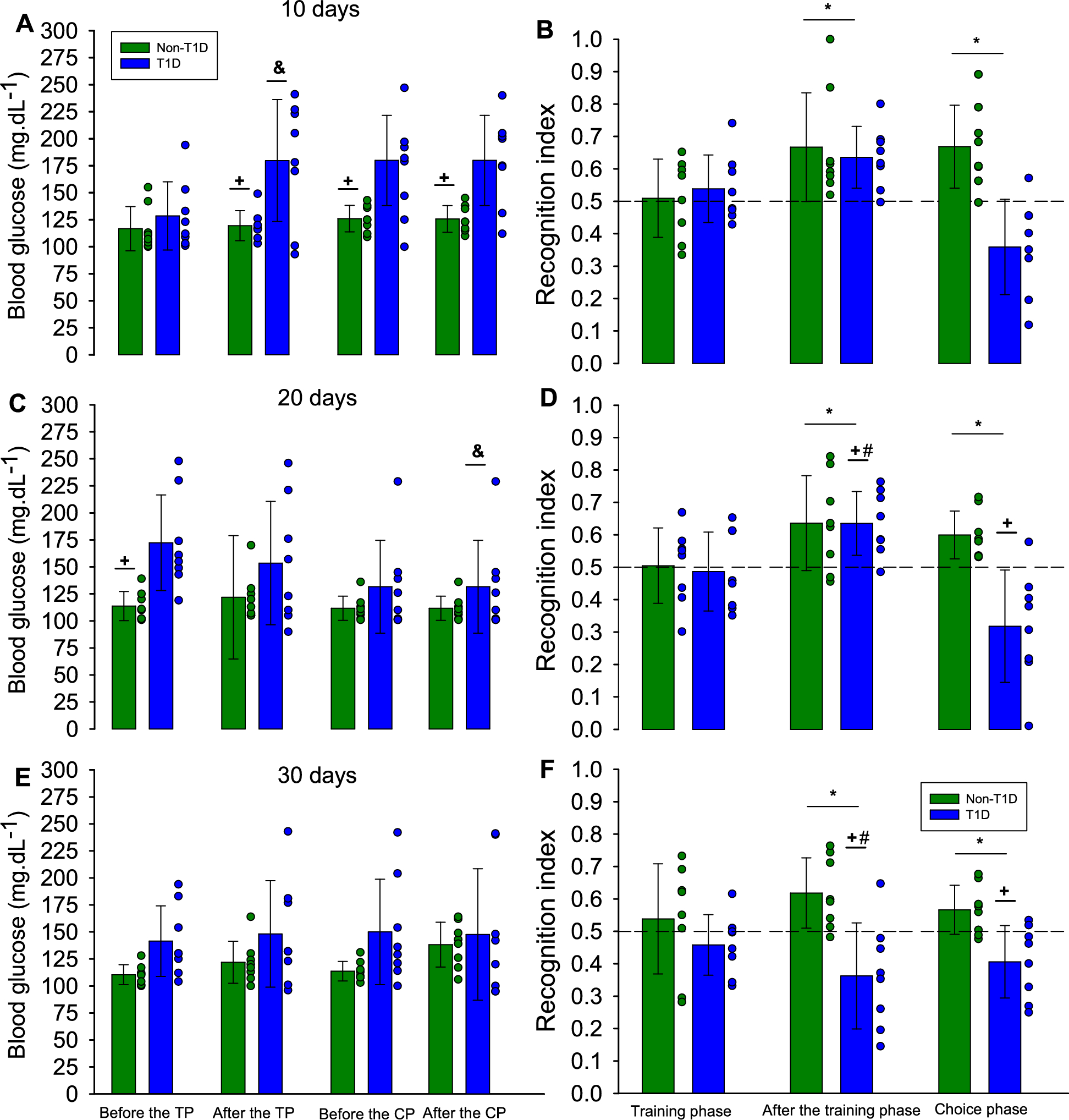
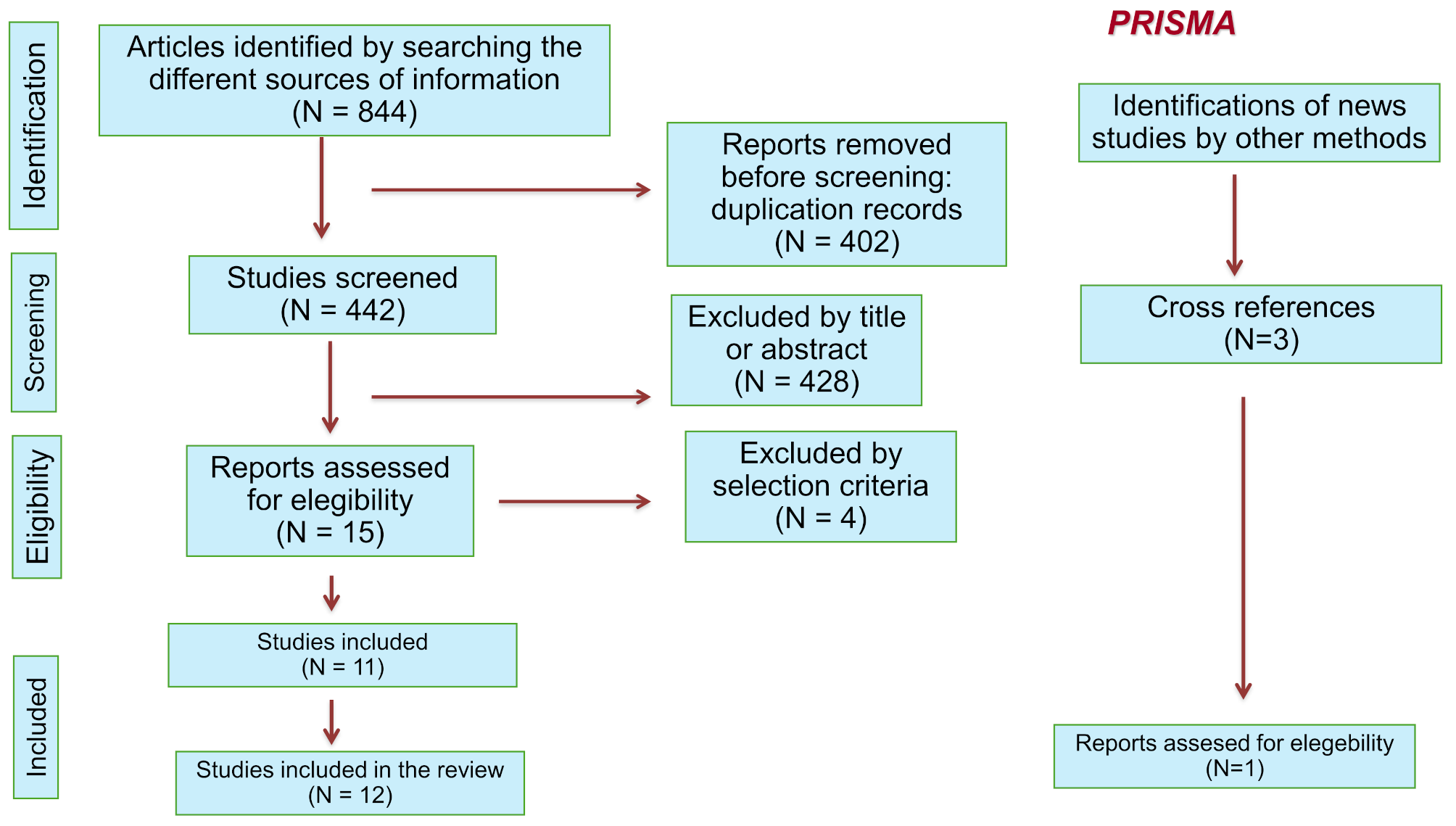
Remember me
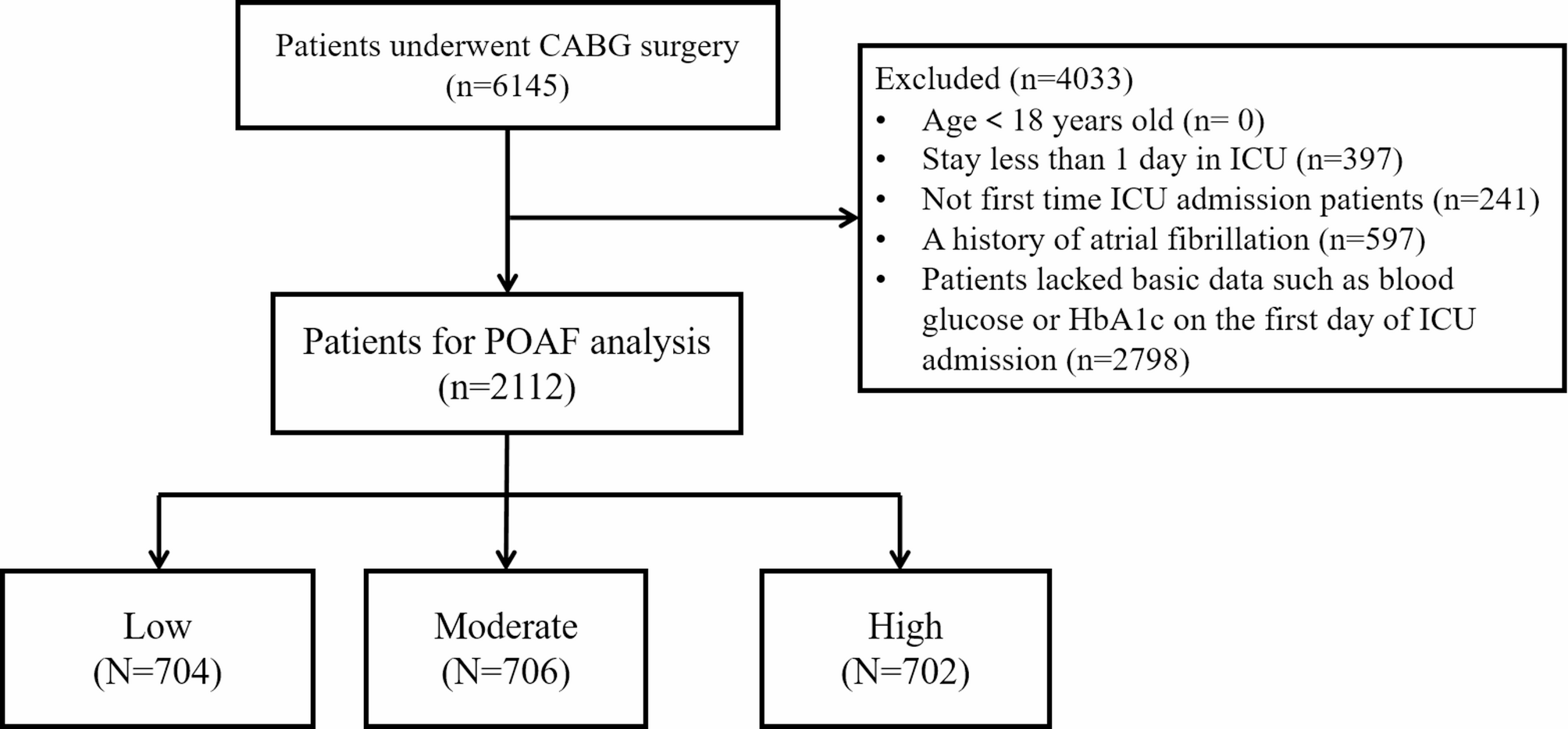
The impact of leptin on migration in 5 mM glucose medium was assessed in murine triple negative metastatic 4T1 and metM-Wntlung breast cancer cell lines. Leptin pretreatment, compared to vehicle, increased migration in both 4T1 and metM-Wntlung cells (Fig. 1A). Leptin also decreased leptin receptor (Ob-R) mRNA expression in 4T1, but not in metM-Wntlung, cells (Fig. 1B), suggesting differential responses to leptin treatment in these cell lines.
Fig. 1
Leptin Effect on Migration of Metastatic Breast Cancer Cells. (A) Both 4T1 and metM-Wntlung cells were pretreated with leptin (300 ng/ml) for 4 days and replated in serum free medium into 8 μm pore transwell inserts and migration assessed. (B) Following leptin treatment or vehicle, mRNA level was measured in 4T1 and metM-Wntlung cells by qPCR. Values are mean ± SEM. Asterisk indicates a significant difference relative to vehicle (p < 0.05) of the same cell type
Given the high metabolic demands of cellular migration, we explored whether AMPK mediates the observed leptin-induced migration. Interestingly, p-AMPK/AMPK was increased in 4T1 cells with leptin treatment (Fig. 2A) but was decreased in metM-Wntlung cells (Fig. 2B), suggesting differential metabolic responses in these two cell lines following leptin treatment. To determine the sufficiency of AMPK signaling to inhibit leptin-induced migration, we treated 4T1 cells with an AMPK inhibitor (compound C) and metM-Wntlung cells with an AMPK activator (AICAR) with or without leptin treatment. While compound C inhibited p-AMPK/AMPK in 4T1 cells (Fig. 2C), no effect of the compound C on leptin-induced migration in 4T1 cells was observed (Fig. 2D), despite increased AMPK activity. Additionally, AMPK activator AICAR reduced AMPK activity (Fig. 2E) but did not affect leptin-induced migration in metM-Wntlung cells (Fig. 2F). Together, these results suggest that although AMPK signaling is differentially regulated, it is not required for leptin-induced migration in either 4T1 or metM-Wntlung cells.
Fig. 2
Leptin Regulation of AMPK Activation in Metastatic Breast Cancer Cells. Representative blots and quantification of p-AMPK and AMPK protein level with vehicle (Veh) or leptin treatment by Western blot with β-actin as a loading control in 4T1 (A) and in metM-Wntlung cells (B). Representative blots of p-AMPK and AMPK protein level with or without the AMPK inhibitor compound C (CC) (1 µM for 2 days) in 4T1 cells (C) and with or without the AMPK activator (AICAR) (1 mM for 2 days) in metM-Wntlung cells (E). (D) The 4T1 cells were pretreated with leptin for 4 days before replating in serum free medium with or without the AMPK inhibitor compound C (CC) (1 µM) into transwell inserts and migration assessed. (F) The metM-Wntlung cells were pretreated with leptin or vehicle for 4 days. During the last 24 h of treatment, cells were incubated with AICAR (1 mM) followed by replating in serum free medium with or without AICAR (1 mM) into transwell inserts and migration assessed. Values are mean ± SEM. Asterisk indicates a significant difference relative to vehicle (p < 0.05) of the same cell type. Groups with different letters are significantly different (p < 0.05) assessed by ANOVA
Leptin is a potent regulator of fatty acid metabolism, including the neutral lipid TAG [21], therefore we assessed TAG levels and FA uptake. Leptin treatment did not change TAG levels or palmitic acid uptake in either cell line (Fig. 3A-B), suggesting that leptin does not alter the contribution of exogenous lipids to cellular lipid pools.
Fig. 3
Leptin Regulation of TAG Level and FA Uptake in Metastatic Breast Cancer Cells. 4T1 and metM-Wntlung cells were treated with vehicle or leptin for 4 days. (A) TAG level was assessed and normalized to total protein per well. (B) Fatty acid uptake was determined following incubation with BODIPY FL-palmitate. The relative BODIPY FL-palmitate uptake is expressed as fluorescence intensity normalized to protein. No significant differences were noted, and values are mean ± SEM
Because FAO has been shown to promote tumor cell migration [22], we investigated FAO’s contribution to the leptin-induced migration in these two cell lines. Relative to vehicle, leptin treatment decreased carnitine palmitoyltransferase 1a (Cpt1a), a rate-limiting enzyme FAO, mRNA level in 4T1 cells (Fig. 4A) but increased Cpt1a mRNA level in metM-Wntlung cells (Fig. 4B). Further, etomoxir, an inhibitor of CPT1, did not affect leptin-induced migration (Fig. 4C-D) in either 4T1 or metM-Wntlung cells, consistent with previous literature utilizing 25 mM glucose [13, 23]. These results suggest that FAO is not required for leptin-induced migration.
Fig. 4
Role of Fatty Acid Oxidation in Leptin-Induced Migration in Metastatic Breast Cancer Cells. Following leptin treatment or vehicle, mRNA levels were measured in 4T1 (A) and metM-Wntlung (B) cells by qPCR. 4T1 (C) and metM-Wntlung (D) cells were pretreated with leptin before replating in serum free medium with or without etomoxir (75 µM) into transwell inserts. Values are mean ± SEM. Asterisk indicates a significant difference relative to vehicle (p < 0.05) of the same cell type. Groups with different letters are significantly different (p < 0.05) assessed by ANOVA
To further explore the role of lipid metabolism in leptin-induced migration, we measured the expression of ATP citrate lyase (Acly) and Fasn following leptin treatment. Fasn mRNA level was suppressed by leptin treatment in 4T1 cells (Fig. 5A) but increased in the metM-Wntlung (Fig. 5B). However, inhibiting fatty acid synthase (TVB-3166) did not suppress leptin-induced migration in metM-Wntlung cells, indicating that fatty acid synthesis is not required for leptin-induced migration (Fig. 5C-D).
Fig. 5
Effect of Inhibiting Fatty Acid Synthase on Leptin-Induced Migration in metM-Wntlung Cells. mRNA level of genes whose protein products are involved in fatty acid synthesis were measured in 4T1 (A) and metM-Wntlung (B) cells after treatment of leptin or vehicle by qPCR. metM-Wntlung cells were pretreated with leptin for 4 days before replating in serum free medium with or without the fatty acid synthase inhibitor TVB-3166 (42 nM) into transwell inserts (C). (D) In each group, the percent increased migration with leptin was calculated by the difference in migration per average of migration in vehicle. Values are mean ± SEM. Asterisk indicates a significant difference relative to vehicle (p < 0.05) of the same cell type. Groups with different letters are significantly different (p < 0.05) assessed by ANOVA. ND indicates no significant difference
Finally, we investigated the contribution of glucose metabolism to leptin-induced migration. Given that in some cancer cells glucose is the predominant source for fatty acids synthesis de novo [24], we measured carbon incorporation from [U]-13C–acetate and [U]-13C–glucose substrates into various fatty acids. First, basal fatty acid synthesis of palmitate from 13C–glucose was similar in 4T1 and the metM-Wntlung cells (4.61 ± 0.1 vs. 4.38 ± 0.15% of total (labeled and unlabeled) palmitate, respectively). Leptin treatment promoted palmitate and stearate synthesis from [U]-13C–glucose in metM-Wntlung but not 4T1 cells (Fig. 6A, C).
The mRNA level of glucose transporter 1 (Glut1), which facilitates glucose transport across the cell membrane, was increased with leptin treatment in both 4T1 and metM-Wntlung cells (Fig. 6B, D). Leptin treatment also increased hexokinase 2 (Hk2), a key glycolytic enzyme, mRNA abundance, in metM-Wntlung cells (Fig. 6D). In addition, intracellular pool sizes of the end products of glycolysis, pyruvate and lactate, were measured after leptin treatment. In basal conditions, pyruvate pools size was 59% greater in metM-Wntlung cells compared to the 4T1 cells (p = 0.005, data not shown), with similar lactate pool sizes (p = 0.79) between the cell lines. Intracellular pool sizes of pyruvate and lactate were induced by leptin treatment in metM-Wntlung but not 4T1 cells (Fig. 6E-F), suggesting that glucose metabolism may be a critical regulator of migration in the metM-Wntlung but not the 4T1 cells.
Fig. 6
Leptin Effects on de novo Fatty Acid Synthesis in Metastatic Breast Cancer Cells. Conversion of glucose or acetate to palmitate or stearate in either vehicle or leptin-treated 4T1 (A) or metM-Wntlung (C) cells is represented as percent of total palmitate or stearate from each 13C substrate. Cells were treated with either vehicle or leptin for 4 days. In separate experiments, either [U]-13C-glucose or [U]-13C-acetate (24 h) incorporation into palmitate or stearate was analyzed using LC-MS/MS. Values are mean ± SEM. mRNA level of genes whose protein products are involved in glucose metabolism were measured in 4T1 (B) and metM-Wntlung (D) cells after treatment of leptin or vehicle by qPCR. Intracellular pools of pyruvate and lactate were measured by GC-MS/MS following leptin treatment in 4T1 (E) and metM-Wntlung (F) cells. Values are mean ± SEM. Asterisk indicates a significant difference relative to vehicle (p < 0.05)
Blocking glycolysis using 2DG (5 mM) potently suppressed leptin-induced migration in metM-Wntlung cells (Fig. 7C-D). Treatment of 4T1 cells with low concentrations (0.3 mM) of 2DG did not affect leptin-induced migration in 4T1 cells (Fig. 7A-B), but higher concentrations (2.5 and 5 mM) of 2DG induced cell death (data not shown), suggesting differential regulation of energy metabolism between the 4T1 and metM-Wntlung cells, with the latter requiring glycolysis for leptin-induced migration.
Fig. 7
Role of Glycolysis in Leptin-Induced Migration in Metastatic Breast Cancer Cells. 4T1 (A, B) and metM-Wntlung (C, D) cells were pretreated with leptin before replating in serum free medium with or without 2-deoxyglucose (2DG) into transwell inserts and migration assessed. The dose of 2DG was 0.3 mM was employed for 4T1 and 5 mM of 2DG was used for metM-Wntlung cells. (B, D) In each group, the percent increase in migration with leptin treatment was calculated by the difference on migration per average of migration in vehicle. Values are mean ± SEM. Asterisk indicates a significant difference relative to vehicle (p < 0.05) of the same cell type. Groups with different letters are significantly different (p < 0.05) assessed by ANOVA. NS indicates no significant difference
Comments (0)