
Remember me




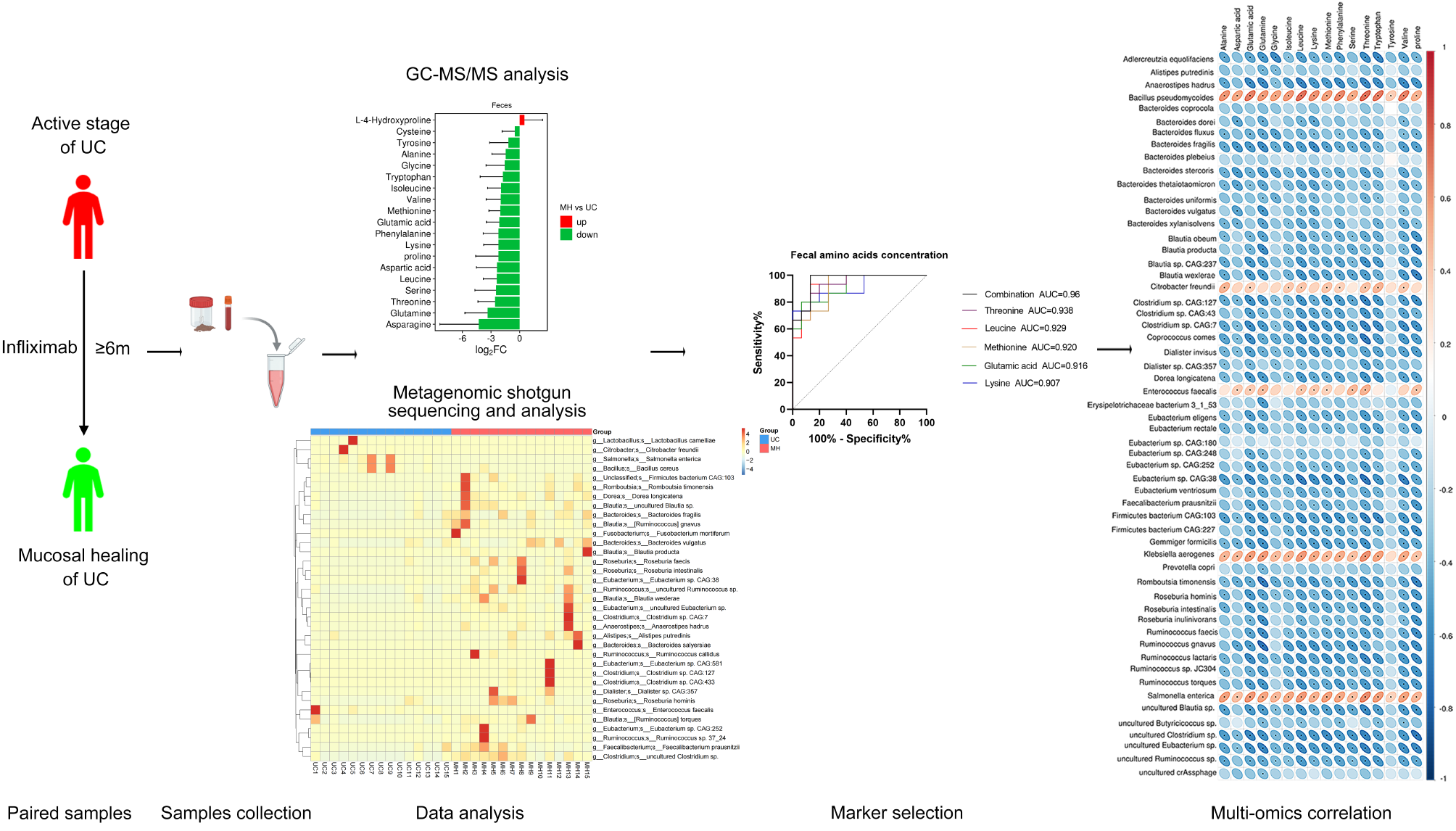
The study population was classified into four groups, and a comparison of clinical features was conducted among these groups. As shown in Table 1 (at the end of the document text file), the median age was similar, ranging from 35 to 40 years. Patients with CHB had a higher proportion of females (75%), whereas patients with CHB and NAFLD had a higher proportion of males (90%). Patients with NAFLD and CHB concurrent with NAFLD exhibited elevated levels of alanine transaminase (ALT), aspartate aminotransferase (AST), Triglyceride(TG), Total cholesterol(TC),γ-glutamyl transpeptadase(GGT), and Uric Acid(UA) compared to the other two groups. These characteristics were comparable between the latter two groups. The groups of CHB and CHB concurrent with NAFLD were significantly positive for HBeAg, HBsAg, and HBV-DNA, although these markers showed no significant differences between the two groups.
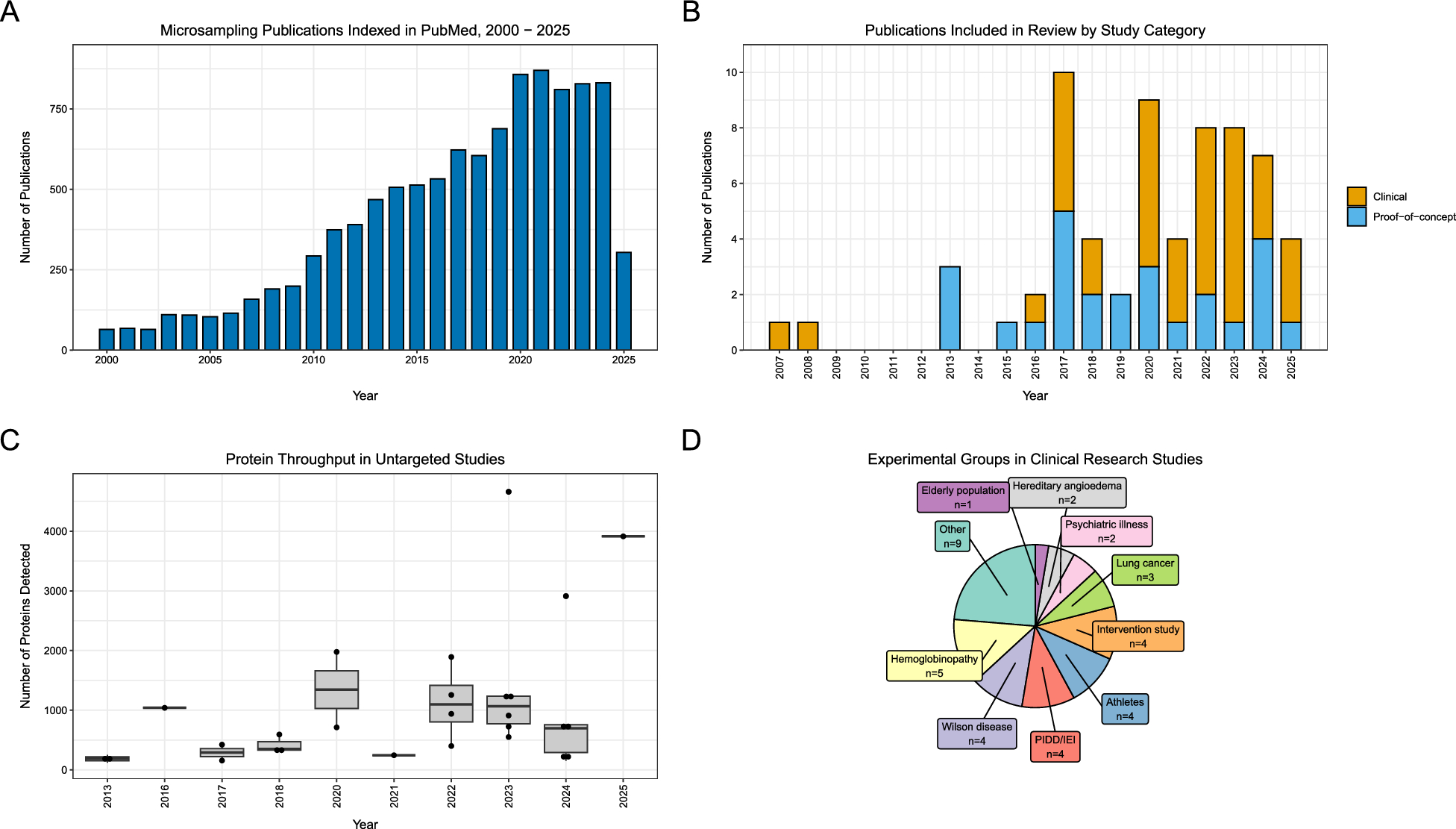
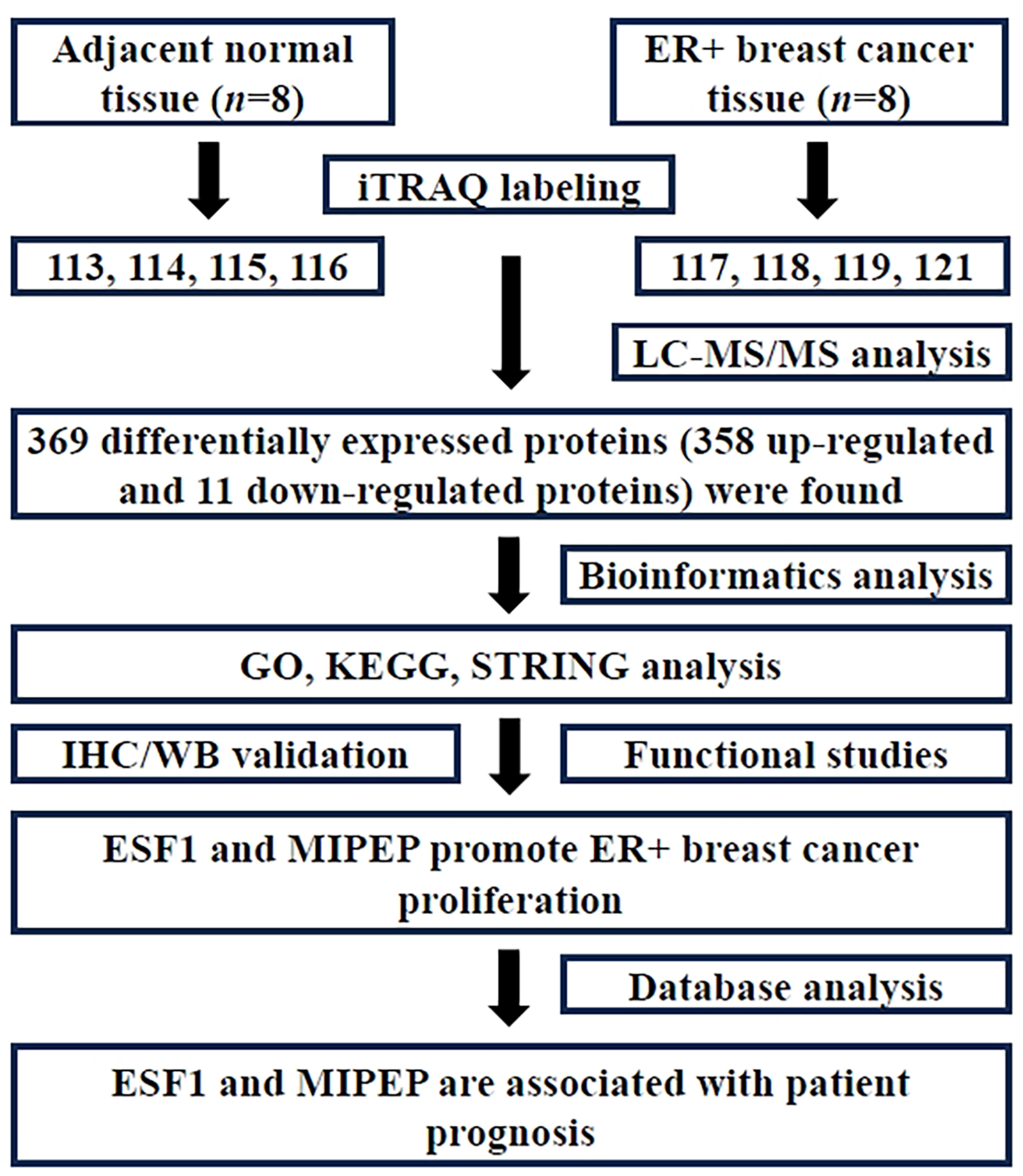
Quantitative protein detectionDIA mass spectrometry was employed to create and analyze a spectral library of human liver proteins derived from 31 subjects, including 8 patients with CHB, 7 patients with NAFLD, 10 patients with CHB and concurrent NAFLD, and 6 healthy individuals. This analysis resulted in the identification of 21,430 peptides and 3,199 proteins (Fig. 1A-B). Compared with the healthy control group, a total of 170 and 100 proteins were found to be differentially expressed in the CHB and NAFLD groups, respectively. Among these DEPs, 51 were upregulated and 129 were downregulated in the CHB group, whereas 80 were upregulated and 20 were downregulated in the group NAFLD. When comparing the CHB group to the group of patients with CHB and NAFLD, a total of 109 proteins were differentially expressed, with 73 being upregulated and 36 downregulated. In comparison to the NAFLD group, a total of 221 proteins were differentially expressed in the group of CHB patients with NAFLD, comprising 22 upregulated and 199 downregulated proteins (Fig. 1C). Among the liver proteins identified, 135 were exclusively detected in CHB samples, 37 were exclusive to NAFLD samples, and only 12 were exclusively present in the samples from patients with CHB and NAFLD. A total of 735 proteins overlapped among the four groups, accounting for 52% of the total quantified proteins (Fig. 1D). To highlight the most significantly dysregulated proteins in patients with CHB and NAFLD compared to patients with only CHB and those with only NAFLD, the top 10 most significant differences in upregulated and downregulated proteins were marked on the volcano plot, using fold change and p-value T-test criteria (Fig. 1E and F).
Fig. 1
Liver protein identification by the DIA for patients in the HC, CHB, NAFLD, CHB concurrent with NAFLD. (A) DIA Experimental Procedure. (B) Quantitative heatmap of total DIA identified proteins for all samples. (C) Histogram of quantitative protein difference results. (D) The Venn diagrams show the numbers of identified proteins and the overlaps of protein identification in the 4 groups. (E) Volcano plot representing the protein abundance changes (groups CHB vs. CHB-NAFLD) (F) Volcano plot representing the protein abundance changes (groups NAFLD vs. CHB-NAFLD)
GO enrichment analysis and protein structural domains analysisBased on these results, we generated GO functional classification maps to compared with standalone CHB, dysregulated proteins in patients with CHB concurrent with NAFLD were associated with processes such as keratinocyte differentiation, epidermal cell differentiation, growth, immunoglobulin complex, and homocysteine metabolic processes. Notably, proteins that were significantly dysregulated between the two groups were primarily enriched in biological processes such as multicellular organismal processes, cellular developmental processes, and cellular amino acid biosynthetic processes (Fig. 2B). In general, protein interactions are often structured in terms of structural domains, and changes in amino acids or modifications within these domains can lead to alterations in key protein functions. The analysis of structural domain enrichment for DEPs indicated that the top five most enriched structural domains were intermediate filament protein, keratin type II head, FAD-dependent oxidoreductase central domain, serpin (serine protease inhibitor), and immunoglobulin V-set domain (Fig. 2A).
The GO enrichment analysis of biological functions related to the proteins that exhibited differential abundance in NAFLD versus CHB concurrent with NAFLD indicated that the CHB concurrent with NAFLD group was enriched in proteins associated with various essential processes. These processes included cellular metabolic processes, organic substance metabolic processes, small molecule metabolic processes, carboxylic acid metabolic processes, and cellular catabolic processes. Functionally, these proteins were primarily associated with antioxidant activity, long-chain fatty acid binding, fatty acid binding, and endopeptidase regulator activity. They were primarily localized in the extracellular organelle, extracellular vesicle, and the extracellular region (Fig. 2C and D).
Fig. 2
(A) Histogram of structural domain analysis of differentially expressed proteins in CHB-NAFLD vs. CHB group. (B) The histograms in gene ontology (GO) analysis of the differentially expressed proteins in CHB-NAFLD vs. CHB. (C) Histogram of structural domain analysis of differentially expressed proteins in CHB-NAFLD vs. NAFLD group. (D) The histograms in GO analysis of the differentially expressed proteins in CHB-NAFLD vs. NAFLD group
KEGG pathway analysisIn addition, KEGG pathway analysis revealed that DEPs in patients were enriched in various biological pathways within KEGG categories. Upregulated proteins were annotated in three major pathways: Staphylococcus aureus infection, the estrogen signaling pathway, and cholesterol metabolism. Conversely, downregulated proteins were annotated in 17 major pathways, with the top three most enriched pathways being valine, leucine and isoleucine degradation, glycine, serine, and threonine metabolism, and butanoate metabolism (Fig. 3A and B). Moreover, propanoate and beta-alanine metabolism, as well as fatty acid degradation, were among the main KEGG pathways related to the differential proteins between the two groups. The differences in various metabolism-related pathways in the livers of patients with CHB concurrent with fatty liver were more pronounced compared to those with standalone CHB, especially with regards to the abnormal activation of cholesterol metabolic pathways, which aligns with the characteristics of fatty liver disease. These changes were primarily associated with the differential enrichment of proteins such as KRT24, SERPINB3, MTCH2, CDSN, SYPL1, AGXT, peroxisome proliferator-activated receptor-gamma coactivator (PGC1-α), and others. Furthermore, after the development of concurrent fatty liver, pathways that were abnormally regulated in CHB alone were instead inhibited. For instance, the glycine, serine, and threonine metabolism pathways were abnormally activated in the CHB group compared to healthy subjects, but they were inhibited in the CHB concurrent with NAFLD group when compared to patients with CHB alone. This inhibition was associated with a decrease in CBS, PSAT1, AGXT, CTH, DMGDH, and SARDH after CHB concurrent with NAFLD (Figure S1). Meanwhile, the interleukin (IL)-17 signaling pathway was activated in the CHB concurrent with NAFLD group compared to the standalone CHB group, primarily associated with the increased expression of S100A7 (Figure S2), known to promote the innate immune response of the host. These findings suggest that the development of CHB may be inhibited after the occurrence of concurrent NAFLD. Interestingly, the CHB concurrent with NAFLD group exhibited the suppression of the HCC pathway compared to the group with CHB alone, and this suppression was associated with elevated LRP6 expression (Figure S3).
Using the KEGG pathway database, KEGG analysis of the NAFLD versus CHB concurrent with NAFLD group revealed 8 major upregulated pathways, including metabolism of xenobiotics by cytochrome P450, IL-17 signaling pathway, and retinol metabolism. In contrast, 36 major pathways were downregulated, including valine, leucine, and isoleucine degradation, beta-alanine metabolism, pentose phosphate pathway, propanoate metabolism, glycolysis/gluconeogenesis, fatty acid degradation, peroxisome proliferator-activated receptor (PPAR) signaling pathway, and fat digestion and absorption. These findings shed light on the inhibition of numerous metabolic pathways in patients with NAFLD in combination with HBV infection, offering new insights into the interaction mechanisms between the two diseases (Fig. 3C and D). Notably, following concurrent HBV infection, pathways that were previously abnormally activated in patients with NAFLD alone were instead inhibited. For instance, glycolysis/gluconeogenesis, pyruvate metabolism, fatty acid degradation, and fat digestion and absorption were abnormally activated in the NAFLD group compared to healthy subjects but were inhibited in the group with CHB and concurrent NAFLD when compared to patients with NAFLD (Figure S4). Key DEPs responsible for these changes included ACADM, ECHS1, ACAT1, EHHADH, and ALDH2. Further confirmation through IHC revealed a significant decrease in positive staining for ECHS1, ACAT1, and ALDH2 after the onset of both diseases.
Fig. 3
(A) KEGG pathway enrichment bubble map of differentially expressed proteins in CHB vs. CHB-NAFLD group. (B) KEGG pathway enrichment bubble map of differentially expressed proteins in NAFLD vs. CHB-NAFLD group. (C) Pathway enrichment butterfly plots of up- and down-regulated differential proteins in the CHB vs. CHB-NAFLD group. (D) Pathway enrichment butterfly plots of up- and down-regulated differential proteins in the NAFLD vs. CHB-NAFLD group
Validation of dysregulated protein expression in patient liver tissuesThe alterations in DEPs were validated at the protein level through IHC, using liver samples from patients (3 from healthy controls, 3 from patients with CHB, 3 from patients with NAFLD, and 3 from patients with CHB + NAFLD). In particular, we assessed three key proteins highlighted in the previously mentioned analyses. Our results confirmed a significant increase in hepatic SERPINB3, ACY1, and MTCH2 positive staining in patients with CHB concurrent with NAFLD compared to patients with CHB alone (Fig. 4A). Conversely, we observed reduced protein levels of hepatic ACAT1, ECHS1, and ALDH2 in patients with CHB and NAFLD compared to patients with NAFLD alone (Fig. 4B). These findings provide valuable insights for future investigations into CHB concurrent with NAFLD, although further study is required to explore the underlying molecular mechanisms.
Fig. 4
(A) Verification of differential proteins between the CHB vs. CHB-NAFLD group by immunohistochemistry. (B) Verification of differential proteins between the NAFLD vs. CHB-NAFLD group by immunohistochemistry (Scale bar: 50 μm)
Validation of dysregulated proteins in mouse modelsTo validate the proteomic results in mouse models, we generated mouse models for HBV, NAFLD, and NAFLD with HBV infection (Fig. 5A). Hepatic hematoxylin and eosin staining revealed reduced hepatocyte volumes, fewer dispersed lipid vacuoles, and compressed liver sinusoids in NAFLD + HBV mice compared to those in NAFLD mice (Fig. 5B). Additionally, we found that the body weight of the NAFLD + HBV group was decreased compared to the NAFLD group, whereas the blood glucose levels did not significantly change in either group (Fig. 5C). Notably, reduced TG and TC secretion levels were identified in NAFLD + HBV mice compared to NAFLD mice (Fig. 5D). Moreover, the serum levels of HBsAg, HBeAg, and HBV-DNA were significantly decreased after 12 weeks of HFD compared to the NAFLD group. Conversely, both serum ALT, AST, triglycerides, and cholesterol in the NAFLD + HBV group were higher than in the NAFLD group, whereas the HBV and control groups exhibited no significant change (Fig. 5E). Furthermore, based on the results of previous analyses, we selected multiple genes involved in hepatic lipogenesis and infection, and examined their expression using qRT-PCR (Fig. 5F).
Fig. 5
(A) Establishment of mouse models of HBV, NAFLD and NAFLD combined with HBV infection. (B) HE staining of liver tissues (C) Changes in body weight of mice over time (D) Serum TG and TC secretion levels in four groups of mice (E) Serum levels of HBsAg, HBeAg and HBV-DNA of mice (F) Validation of differential gene expression by qRT-PCR
Comments (0)