
Remember me

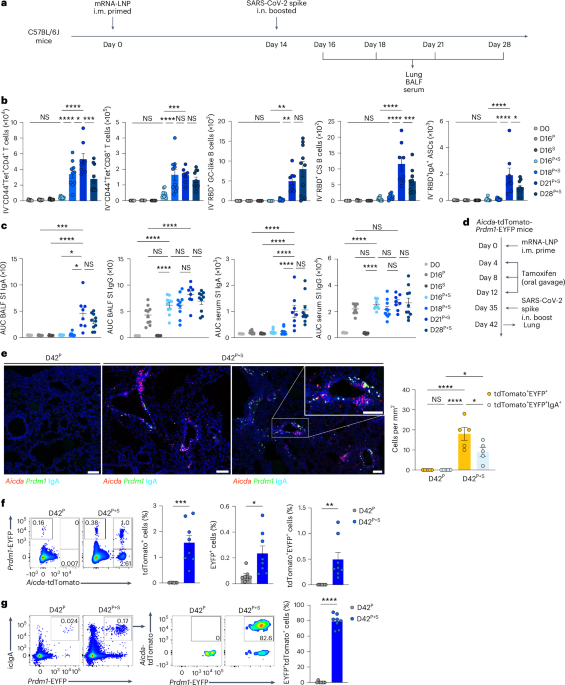

We first assessed the time course of accumulation of pulmonary antigen-specific T cells and B cells during the ‘prime and spike’ (hereafter P + S) vaccine approach7, in which C57BL/6J mice were injected intramuscularly (i.m.) with a 1 μg mRNA–LNP vaccine encoding the full-length SARS-CoV-2 spike protein (Pfizer/BioNTech BNT162b2) at day 0, followed by boosting i.n. with an unadjuvanted 1 μg SARS-CoV-2 spike protein at day 14 (Fig. 1a). CD45 labeling intravenously (i.v.) and spike-specific major histocompatibility complex class I (MHC-I) and MHC-II tetramers indicated a notable increase in the number of spike-specific, tissue-resident CD4+ and CD8+ T cells in the lungs at day 18 after primary vaccination, with a peak at day 21 post-priming i.m. (Fig. 1b and Extended Data Fig. 1a). Next, we identified antigen-specific, tissue-resident B cells and IgA+ antibody-secreting cells (ASCs) using SARS-CoV-2 receptor-binding domain (RBD)-specific tetramers and intracellular staining with antibodies specific for IgA and BLIMP1, a transcription factor essential for plasma cell generation14. At day 7 after boosting i.n., we detected a substantial increase in tissue-resident, RBD-specific CD38−GL7+ germinal center (GC)-like and CD38+GL7−IgM−IgD− class-switched B cells, and icIgA+BLIMP1+ ASCs in the lungs (Fig. 1b and Extended Data Fig. 1b). Notably, only mice that received the booster i.n. showed an increase in SARS-CoV-2 S1 subunit-specific IgA amount in the bronchoalveolar lavage fluid (BALF) and serum (Fig. 1c). Although priming with mRNA–LNP vaccine alone elicited S1-specific IgG in the serum, BALF S1-specific IgG levels increased at day 21 post-priming i.m. (Fig. 1c). Thus, unadjuvanted spike protein delivery i.n. rapidly triggered strong antigen-specific CD4+ and CD8+ T cell and IgA responses in the lung.
Fig. 1: Unadjuvanted nasal booster elicits generation of antigen-specific IgA+ plasma cells at the respiratory mucosa.
a, Schematic of the experimental setup showing C57BL/6J mice immunized i.m. with mRNA–LNPs encoding the full-length SARS-CoV-2 spike protein at day 0, i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 14, followed by lung, BALF and serum collection at days 16, 18, 21 and 28 post-priming i.m. b, Number of extravascular CD44+Tet+CD4+ T cells, CD44+Tet+CD8+ T cells, RBD+CD38−GL7+ GC-like B cells, RBD+CD38+GL7−IgM−IgD− class-switched (CS) B cells and RBD+IgA+ ASCs in the lungs of naive C57BL/6J mice (day 0 (D0), n = 9); intramuscular prime-only mice (D16P, n = 10) and intranasal booster-only mice (D16S, n = 10) at day 16 post-priming i.m. and P + S mice at day 16 (D16P+S, n = 10), day 18 (D18P+6, n = 10), day 21 (D21P+S, n = 8) and day 28 (D128P, n = 10) post-priming i.m., analyzed by flow cytometry as in a. c, ELISA measurements of SARS-CoV-2 spike S1 subunit-specific IgA and IgG in the BALF and serum from mice as in b. AUC, area under the curve. d, Schematic of experiment showing Aicda-tdTomato-Prdm1-EYFP mice immunized i.m. with mRNA–LNPs at day 0, treated with 10 mg of tamoxifen via oral gavage at days 4, 8 and 12, boosted i.n. with a spike protein at day 35, followed by lung collection at day 42 post-priming. e, Immunofluorescence of IgA staining (left) and quantification (right) of colocalization of tdTomato+EYFP+ and tdTomato+EYFP+IgA+ cells in the lungs of intramuscular prime-only (D42P, n = 5) or P + S (D42P+S, n = 5) Aicda-tdTomato-Prdm1-EYFP mice at day 42 post-priming i.m. (day 7 post-boosting i.n.) as in d. Scale bar, 100 μm. f, Representative flow cytometry plots and frequency of EYFP+, tdTomato+ and tdTomato+EYFP+ cells in the lungs of intramuscular prime-only (D42P, n = 7) and P + S (D42P+S, n = 8) Aicda-tdTomato-Prdm1-EYFP mice analyzed by flow cytometry at day 42 post-priming i.m. as in d. g, Representative flow cytometry plots and frequency of tdTomato+ cells among EYFP+IgA+ cells in the lungs of mice as in f (mean ± s.e.m.). Statistical significance was calculated using one-way ANOVA (b,c,e) and unpaired Student’s t-test (f,g). Tukey’s multiple comparisons (b,c,e) were performed: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. NS, not significant. Data were pooled from three (b,c) independent experiments or represented (e–g) two independent experiments. Values indicated as zero represent the absence of detectable cells (b,e–g).
To determine whether direct delivery of unadjuvanted nasal boosters into the lungs was required for eliciting mucosal recall responses, we compared antigen-specific T and B cell immunity in C57BL/6J mice primed with 1μg mRNA–LNPs at day 0, followed by boosting at day 14 with either 1 μg of mRNA–LNPs i.m. or 1 μg of SARS-CoV-2 spike protein i.n., intratracheally (i.t.), or intraperitoneally (i.p.) (Extended Data Fig. 2a). Boosting i.t. with an unadjuvanted SARS-CoV-2 spike protein substantially increased the number of spike-specific CD4+ and CD8+ T cells, as well as RBD-specific IgA+ ASCs in the lungs compared with boosting i.m. with mRNA–LNPs and i.p. with spike protein (Extended Data Fig. 2b). Boosting i.m. with mRNA–LNPs and systemic boosting i.p. with SARS-CoV-2 spike protein did not induce RBD-specific IgA+ ASCs in the lung (Extended Data Fig. 2c). Only mice boosted i.n. or i.t. showed an increase in S1-specific IgA levels in BALF, whereas serum S1-specific IgG levels were higher in mice boosted i.m. with mRNA–LNPs at day 14 post-priming i.m. (Extended Data Fig. 2d). Thus, antigen delivery directly into the respiratory mucosa was necessary to generate robust local CD4+ and CD8+ T cell and IgA responses. The numbers of spike-specific CD4+ and CD8+ T cells, RBD-specific IgA+ ASCs and S1-specific IgA and IgG levels in BALF and serum were comparable between female and male mice boosted i.n. with spike protein at day 14 post-priming i.m. (Extended Data Fig. 2e–g), indicating that an unadjuvanted intranasal protein booster induced robust immune responses in respiratory mucosa independent of sex.
Next, we investigated the localization of IgA+ plasma cells and memory B cells in Aicda-ERT2-Cre-Rosa26-tdTomato Prdm1-enhanced yellow fluorescent protein (EYFP) mice treated with tamoxifen at days 4, 8 and 12 after immunization i.m. with mRNA–LNPs, followed by boosting i.n. with SARS-CoV-2 spike protein at day 35 (Fig. 1d), an experimental setup in which activated B cells are permanently tdTomato+ and primed B cell-derived plasma cells are tdTomato+EYFP+. Although not all activated B cells are fate mapped in the Aicda-ERT2-Cre mice15, we observed predominantly tdTomato+ primed B cell (red) clusters, along with a mixture of EYFP+ plasma cells (green), tdTomato+EYFP+ primed B cell-derived plasma cells (yellow) and tdTomato+EYFP+IgA+ plasma cells (white) derived from B cells primed i.m. near the blood vessels and airways in the lung parenchyma at day 42 post-priming i.m. (Fig. 1e). All these B cell populations were absent in intramuscular prime-only mice (Fig. 1e). Flow cytometry analysis of whole lung tissue also indicated a substantial increase in frequency of tdTomato+, EYFP+ and tdTomato+EYFP+ cells in the lungs of i.n. boosted compared with intramuscular prime-only Aicda-ERT2-Cre-Rosa26-tdTomato Prdm1-EYFP mice at day 42 post-priming i.m. (Fig. 1f). More than 80% of IgA+ plasma cells in the mice boosted i.n. were tdTomato+ (Fig. 1g), suggesting that IgA-producing plasma cells mainly originated from activated B cells formed during priming i.m. Thus, boosting i.n. with an unadjuvanted spike protein induced IgA+ plasma cell development in the respiratory mucosa.
Mucosal IgA responses originate from circulating memory cellsTo identify the source of antigen-specific mucosal responses, CD45.2 mice primed i.m. with mRNA–LNPs at day 0 were surgically paired with CD45.1 naive mice at day 14 to establish parabiotic pairs in which the circulatory systems are known to equilibrate within 2 weeks16. At week 2 post-surgery, either the CD45.1 naive or the CD45.2 i.m. primed mice of the parabiosis pair received an unadjuvanted SARS-CoV-2 spike protein i.n. (Fig. 2a). At day 14 post-boosting i.n., the frequency of host-derived cells among intravenously (i.v.) CD45-labeled cells in the lung of each parabiont was comparable (Fig. 2b), indicating that the circulatory immune cells had reached an equilibrium. The number of spike-specific CD4+ and CD8+ T cells was markedly higher in the lung of i.n. boosted parabiont compared with the non-i.n. boosted partner parabiont (Fig. 2c,e). Notably, >90% of the spike-specific CD4+ and CD8+ T cells originated from CD45.2+ T cells derived from the i.m. primed parabiont (Fig. 2d,f), suggesting that mucosal recall responses were mostly derived from circulating memory T cells rather than naive T cells after boosting i.n. RBD-specific IgA+ ASCs were predominantly present in the lung of the i.n. boosted parabiont and most of them were CD45.2+ (Fig. 2g,h), suggesting that they originated from the i.m. primed parabiont. BALF S1-specific IgA was markedly elevated in the i.n. boosted parabionts, but undetectable in the non-i.n. boosted parabiont (Fig. 2i), indicating local production and secretion of IgA. S1-specific IgG levels were higher in the BALF of the i.n. boosted parabionts compared with the non-i.n. boosted parabiont (in which the BALF S1 IgG was still highly detectable) (Fig. 2i), suggesting that BALF S1-specific IgG was primarily derived from systemic sources after boosting i.n. Thus, circulating immune cells were sufficient to elicit antigen-specific antibody responses in the lung after boosting i.n. and local mucosal T cell and IgA responses that developed after this nasal booster with an unadjuvanted SARS-CoV-2 spike protein originated from those primed during the intramuscular mRNA–LNP vaccination.
Fig. 2: Circulating memory cells are the source for mucosal recall responses.
a, Schematic of the experimental design showing CD45.2 C57BL/6J mice primed i.m. with mRNA–LNPs at day 0 surgically paired with CD45.1 naive mice at day 14, followed by an intranasal boost of either the CD45.1 naive or the CD45.2 primed mice of the parabiotic pair with SARS-CoV-2 spike protein at day 14 after surgery, and analysis of lungs and BALF at day 14 after booster i.n. b, Representative flow cytometry plots and frequency of host-derived cells among intravenous CD45+ cells in the lungs of a parabiosis pair boosted i.n. in either CD45.1 (n = 17) or CD45.2 (n = 15) mice analyzed by flow cytometry as in a. c–f, Representative flow cytometry plots (left) and number (right) of extravascular IV−CD44+Tet+CD4+ T cells (c) and IV−CD44+Tet+CD8+ T cells (e) and frequency of CD45.2+ cells among extravascular IV−CD44+Tet+CD4+ T cells (d) and IV−CD44+Tet+CD8+ T cells (f) in the lungs of CD45.1 and CD45.2 mice of a parabiosis pair as in b. g, Representative flow cytometry plots and extravascular RBD+IgA+ ASC numbers in the lungs of CD45.1 (n = 5) and CD45.2 (n = 5) mice of a parabiosis pair as in a. h, Frequency of CD45.2+ cells among extravascular IV−RBD+IgA+ ASCs in the lungs of CD45.1 and CD45.2 mice of a parabiosis pair as in g. i, ELISA measurements of SARS-CoV-2 S1-specific IgA and IgG in the BALF of CD45.1 and CD45.2 mice of a parabiosis pair as in b. Data are the mean ± s.e.m. The statistical significance was calculated using one-way ANOVA (b,i) or nonparametric Kruskal–Wallis test (c,e,g). Tukey’s multiple comparisons (b,i) and Dunn’s multiple comparisons (c,e,g) were performed: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Data were pooled from four (b–f) independent experiments or represent (g,h) an experiment. Values shown as zero indicate the absence of detectable cells (c,e,g).
Lymph node memory B cells are the main source of mucosal IgA responsesIntramuscular vaccination with mRNA–LNPs induces a robust GC response in the draining lymph nodes in both mice and humans within 1–3 weeks17,18,19,20. Consistently, the mRNA–LNP vaccination i.m. notably induced antigen-specific GC B cell responses in the inguinal lymph nodes, but not in the spleen of C57BL/6J mice at day 7 after immunization i.m. (Extended Data Fig. 3a). As the origin of memory B cells that seed the antigen-specific IgA+ B cells in the lung mucosal tissues remains unknown, we investigated whether lymph nodes contained B cells that could be induced to secrete IgA on boosting i.n. C57BL/6J mice primed i.m. with mRNA–LNPs (day 0) were treated with phosphate-buffered saline (PBS) or the S1PR1 agonist FTY720, which inhibits lymphocyte egress from the secondary lymphoid tissues, every other day for a week starting at days 0, 2 or 4 post-boosting i.n. (day 14) with an unadjuvanted SARS-CoV-2 spike protein (Fig. 3a). Lung RBD-specific IgA+ ASCs, CD38−GL7+ GC-like B cells and CD38+GL7−IgM−IgD− class-switched B cells disappeared almost entirely in the i.n. boosted mice treated with FTY720 at day 0 after an intranasal booster, but were detected in i.n. boosted mice treated with FTY720 at days 2 and 4 with similar levels to i.n. boosted mice not treated with FTY720 (Fig. 3b and Extended Data Fig. 3b,c). BALF S1-specific IgA levels were almost undetectable in i.n. boosted, day 0, FTY720-treated mice, but remained unaffected in i.n. boosted, day 2 and 4 FTY720-treated mice (Fig. 3c). S1-specific BALF IgG and serum IgA levels were substantially reduced, whereas serum S1-specific IgG was minimally affected in i.n. boosted, day 0, FTY720-treated mice compared with i.n. boosted, PBS-treated mice (Fig. 3c). These observations indicated that IgA+ B cells in the lung originated in the lymph nodes and exited within 2 d of boosting i.n. In addition, the number of i.n. boosted, spike-specific, lung CD4+ and CD8+ T cells were markedly reduced in day 0, but not day 2 or 4, FTY720-treated mice, compared with PBS-treated mice (Extended Data Fig. 3d), suggesting that lymph node egress was required for the induction of mucosal T cell and antibody responses.
Fig. 3: Lymph node egress is necessary to generate anamnestic mucosal IgA responses.
a, Schematic of the experimental setup showing C57BL/6J mice i.m. primed with mRNA–LNPs at day 0, i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 14 post-priming, followed by treatment i.p. with PBS or FTY720 every other day, starting at day 14 (treatment 1), 16 (treatment 2) or 18 (treatment 3) after intramuscular priming, and lung, BALF and serum collection at day 21 after an intramuscular boost. b, Representative flow cytometry plots and number of extravascular total and RBD-specific IgA+ ASCs in the lungs of naive C57BL/6J mice (D0, n = 12); P + S mice treated with PBS (D21P+S, n = 13) or P + S mice treated with FTY720 at day 14 (D21T1, n = 10); day 16 (D21T2, n = 9); P + S day 18 (D21T3, n = 9) analyzed by flow cytometry at day 21 after intranasal priming as in a. c, ELISA measurements of SARS-CoV-2 S1-specific IgA and IgG in the BALF and serum from mice as in b. Data are the mean ± s.e.m. The statistical significance was calculated using one-way ANOVA (b,c). Tukey’s multiple comparisons were performed: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Data were pooled from three independent experiments (b,c). Values indicated as zero show the absence of detectable cells (b).
To examine whether mucosal B cells originated from primary GC-derived cells, S1pr2-ERT2-Cre-Rosa26-tdTomato mice, which allow fate mapping of tdTomato+ GC B cells after tamoxifen treatment, were i.m. primed with mRNA–LNPs at day 0 and i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 32 after intramuscular priming. Intramuscularly primed S1pr2-ERT2-Cre-Rosa26-tdTomato mice were i.p. treated with 4-hydroxytamoxifen at days 6, 7, 8, 9 and 10 after priming i.m. or at days 2, 3, 4, 5 and 6 after boosting i.n. (Extended Data Fig. 4a). At day 39 post-primary immunization, >90% of lung RBD-specific B cells and IgA+ ASCs were tdTomato− in mice treated with tamoxifen after boosting i.n. (Extended Data Fig. 4b). By contrast, >60% of them were tdTomato+ in mice given tamoxifen after priming i.m. (Extended Data Fig. 4b), indicating that lung mucosa IgA recall responses were derived from GC B cells formed during primary immunization i.m., not post-boosting i.n. Thus, lymph node-resident memory B cells formed during primary vaccination have a critical role in mucosal recall IgA responses.
CXCR3-dependent signaling recruits memory lymphocytesTo identify early changes in immune cell populations on boosting i.n., we performed single-cell RNA sequencing (scRNA-seq) of extravascular lung CD45+ cells from P + S C57BL/6J mice at day 2 after intranasal boosting. Among the 16 clusters identified, i.n. boosted mice had increased subsets of Ifng+Gzmb+ natural killer (NK) cells (cluster C6) and memory or activated Cd44+Cxcr3+Cd4+ and Cd44+Cxcr3+Cd8+ T cells (C3 and C5), as well as Cxcl9+Cxcl10+ monocyte-derived dendritic cells (moDCs, C1) and Ly6g−Cxcr2+ and Ly6g−Ccl3+ neutrophils (C8 and C9) compared with i.m. primed mice (Extended Data Fig. 5a–d), suggesting that the intranasal booster induced the recruitment of NK cells, memory T (TM) cells and innate immune cells to the lungs. Gene Ontology (GO) analysis showed that genes upregulated at day 2 after boosting i.n. compared with priming i.m. only were associated with inflammatory response, antigen processing and presentation and defense response to the virus (Extended Data Fig. 5e). Also, effector or memory CD4+ T cell clusters expressed Cd40lg and Tgfb1, which are essential for T cell-dependent, IgA class-switch recombination (CSR)21,22,23,24,25 (Extended Data Fig. 5f). Thus, an unadjuvanted spike intranasal booster shaped immune-stimulating microenvironmental niches to elicit anamnestic immunity in the lung.
To address whether early changes in the lung mucosa innate cell populations induced recruitment of memory lymphocytes to the lung, we measured the production of chemokines in BALF of P + S C57BL/6 mice at days 2, 4 and 7 after boosting i.n. with SARS-CoV-2 spike protein by ELISA (Fig. 4a). Expression of CXCL9 and CXCL10, which are strong chemoattractants for TM cells and memory B (BM) cells26, was markedly elevated at day 2 after boosting i.n. and rapidly decreased at day 4 after boosting i.n. (Fig. 4b). At day 7 after boosting i.n., lung Tet+PD-1+CD4+ T cells, Tet+PD-1+CD8+ T cells and RBD+ B cells had much higher expression of CXCR3 compared with lung tetramer− or circulating i.v. CD45-labeled CD4+ T cells, CD8+ T cells and B cells analyzed by flow cytometry (Fig. 4c). To test the role of CXCR3–CXCL9 and CXC3–CXCL10 signaling in the recruitment of CD44+Tet+ TM cells and RBD+CD38+IgD− BM cells to the lung, C57BL/6J mice i.m. primed with mRNA–LNPs were injected with blocking antibodies specific for CXCR3, CXCL9 and CXCL10 (to induce complete blockade of CXCR3–CXCL9-CXCL10 signaling) immediately after boosting i.n. with an unadjuvanted SARS-CoV-2 spike protein (Fig. 4d). We found a substantial reduction in CD44+Tet+CD4+ T cells, CD44+Tet+CD8+ T cells, RBD-specific CD38−GL7+ GC-like B cells, RBD-specific CD38+GL7−IgM−IgD− class-switched B cells and RBD-specific IgA+ ASCs in the lungs (Fig. 4e), as well as S1-specific IgA in BALF and serum, but not serum S1-specific IgG (Fig. 4f) in mice treated with CXCR3 + CXCL9 + CXCL10-blocking antibodies compared with mice treated with PBS. These results indicated that CXCR3–CXCL9 and CXCR3–CXCL10 signaling recruited TM cells and BM cells required for the intranasal booster-mediated responses.
Fig. 4: CXCR3–CXCL9 or CXCR3–CXCL10 signaling is crucial for memory T cell and B cell recruitment into the lung on mucosal boosting.
a, Schematic of the experimental setup showing C57BL/6J mice immunized i.m. with mRNA–LNPs at day 0, i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 14, followed by lung and BALF collection at days 16, 18 and 21 after priming i.m. b, ELISA measurements of CXCL9 and CXCL10 concentrations in the BALF from naive C57BL/6J mice (D0, n = 10); intramuscular prime-only (D16P, n = 9) and intranasal booster-only (D16S, n = 10) mice at day 16; and P + S mice at day 16 (D16P+S, n = 15), day 18 (D18P+S, n = 10) and day 21 (D121P+S, n = 9) as in a. Detection limits, CXCL9 (0.23 pg ml−1) and CXCL10 (0.63 pg ml−1). c, Representative histogram of CXCR3 expression in extravascular IV−PD-1+Tet+ and IV−PD-1−Tet− and circulating IV+CD4+ T and CD8+ T cells and extravascular IV−RBD+ and IV−RBD− and circulating IV+ B cells in the lungs of P + S mice at day 21 post-priming. d, Schematic of the experimental design showing C57BL/6J mice immunized i.m. with mRNA–LNPs at day 0, i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 14, followed by being i.p. treated with PBS or CXCR3 + CXCL9 + CXCL10 neutralizing antibodies at days 14, 16, 18 and 20 after intramuscular priming and lungs, BALF and serum were analyzed at day 21 after priming i.m. e, Number of extravascular IV−CD44+Tet+CD4+ T cells, IV−CD44+Tet+CD8+ T cells, IV−RBD+CD38−GL7+ GC-like B cells, IV−RBD+CD38+GL7−IgM−IgD− class-switched B cells and IV−RBD+IgA+ ASCs in the lung of naive mice (D0, n = 7) and P + S mice treated with PBS (D21PBS, n = 9) or CXCR3 + CXCL9 + CXCL10-neutralizing antibodies (D21Ab, n = 13) at day 21 after priming i.m. as in d analyzed by flow cytometry. f, ELISA measurements of SARS-CoV-2 spike S1-specific IgA and IgG in the BALF and serum from mice as in e. Data are the mean ± s.e.m. The statistical significance was calculated using one-way ANOVA (b,e,f). Tukey’s multiple comparisons were performed: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Data were pooled from three (b) or two independent (e,f) experiments or represent (c) two independent experiments. Values shown as zero indicate the absence of detectable cells (b,e).
Endotoxin-free nasal booster induces mucosal recall responsesLipopolysaccharide (LPS) triggers innate immune activation through the activation of the toll-like receptor-4, resulting in the accumulation of proinflammatory innate cells and cytokines in the lungs27,28. To rule out the possibility that LPS contamination during the preparation of the SARS-CoV-2 spike protein served as an adjuvant when using the spike protein as an intranasal booster, we measured the expression of several proinflammatory cytokines in the BALF of mice i.n. administered with the SARS-CoV-2 spike protein, and found that the expression of BALF interleukin-1α (IL-1α), IL-1β, IL-17A, tumor necrosis factor (TNF) and granulocyte–colony-stimulating factor (G-CSF) proteins was comparable at day 2 after administration between intranasal spike protein-treated and naive mice (Extended Data Fig. 6a). To assess whether LPS would activate the lung-resident innate immune cells when i.n. administered, C57BL/6J mice were i.n. treated with SARS-CoV-2 spike protein along with 0.4 pg and 1,000 pg of LPS. Intranasal administration of spike protein with even a trace amount of LPS (0.4 pg) was sufficient to notably increase the number of Ly6C+ moDCs and CD11b+Ly6G+ neutrophils in the lung mucosa at day 2 after treatment, whereas spike protein alone did not induce any notable changes compared with naive mice (Extended Data Fig. 6b), suggesting that the spike protein that we used in the intranasal booster did not contain innate immune stimulants. Next, mice primed i.m. with mRNA–LNPs were boosted i.n. with spike protein either before or after LPS removal using an endotoxin removal kit (Extended Data Fig. 6c). We observed no substantial differences in the number of spike-specific T cells and RBD-specific B cells in the lung mucosa (Extended Data Fig. 6d), as well as S1 IgA and IgG in the BALF and serum in mice i.n. boosted with either type of spike protein (Extended Data Fig. 6e). Collectively, these data indicated that robust cellular and humoral immune responses in the lung mucosa after an unadjuvanted intranasal booster administration were not driven by LPS contamination and that the spike protein itself could be used as a safe intranasal vaccine booster.
Pre-existing CD4+ T cells serve as natural adjuvantsCD4+ T cells provide local help for CD8+ T cell and B cell response against virus infection29,30, but their role in recall immunity after mucosal boosting remains unclear. To examine the role of pre-existing CD4+ T cells in lung immune cell accumulation within 2 d after boosting i.n., mRNA–LNP i.m. primed C57BL/6J mice were i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 15, followed by intranasal treatment of PBS or a CD4-depleting antibody at days 14 and 16 post-primary immunization (Extended Data Fig. 7a). At day 2 after boosting i.n. with spike protein, mice treated with an intranasal CD4 antibody had an almost complete loss of BALF CXCL9 and CXCL10 as well as lung CD44+Tet+CD8+ T cells and Ly6C+ moDCs compared with mice treated i.n. with PBS (Extended Data Fig. 7b–d), indicating an essential role of pre-existing CD4+ T cells in recruiting and activating the CD8+ T and innate immune cells into the lung. To test whether CD4+ T cells were also central for mucosal IgA recall response, C57BL/6J mice i.m. primed with mRNA–LNPs were i.n. boosted with an unadjuvanted SARS-CoV-2 spike protein at day 14 after priming i.m., followed by intranasal treatment with a CD4-depleting antibody at days 0, 2 and 4 after boosting i.n. (Fig. 5a). CD4 depletion led to a marked reduction in the number of RBD-specific, CD38−GL7+ GC-like B cells and IgA+ ASCs in the lung mucosa (Fig. 5b), as well as S1-specific IgA and IgG in the BALF, whereas systemic S1-specific IgG levels were unchanged compared with PBS treatment (Fig. 5c). Thus, CD4+ TM cells were indispensable for anamnestic IgA responses in the lung mucosa.
Fig. 5: CD4+ T cells are indispensable for generating IgA recall response and providing protective mucosal immunity.
a, Schematic of the experimental setup showing C57BL/6J mice primed i.m. with mRNA–LNPs at day 0, i.n. boosted with an unadjuvanted recombinant SARS-CoV-2 spike protein at day 14, followed by intranasal treatment with PBS or a CD4-depleting antibody at days 14, 16 and 18 after intramuscular priming and lung, BALF and serum collection at day 21 post-priming i.m. b, Representative flow cytometry plots and number of extravascular IV−RBD+CD38−GL7+ GC-like B cells and IV−RBD+IgA+ ASCs in the lungs of naive C57BL/6J mice (D0, n = 8); P + S mice treated with PBS (D21PBS, n = 10) or a CD4-depleting antibody (D21Ab, n = 9), analyzed by flow cytometry at day 21 as in a. c, ELISA measurements of SARS-CoV-2 S1-specific IgA and IgG in the BALF and serum from mice as in b. d, Weight loss and survival at days 0–14 post-infection with SARS-CoV-2 (WA1/2020) in naive K18-hACE2 mice (n = 6), prime-only mice (P, n = 9) and P + S mice treated with PBS (P + SPBS, n = 14) or CD4-depleting antibody (P + SAb, n = 12) either i.n. or i.p. at days 14, 16, 18, 20, 23 and 26 post-priming i.m. and infected with SARS-CoV-2 (WA1/2020) at day 42 post-priming i.m. e, Lung viral titer in naive mice (n = 10); intramuscular prime-only mice (P, n = 8) and P + S mice treated with PBS (P + SPBS, n = 10) or a CD4-depleting antibody (P + SAb, n = 10) as in d at day 2 post-infection with SARS-CoV-2 (WA1/2020) measured by plaque assay. Data are the mean ± s.e.m. The statistical significance was calculated using one-way ANOVA (b, c), log(rank) Mantel–Cox test (d) or nonparametric Kruskal–Wallis test (e). Tukey’s multiple comparisons (b, c), Bonferroni’s multiple comparisons (d) and Dunn’s multiple comparisons (e) were performed: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Data were pooled from two (b–e) independent experiments. Values indicated as zero represent the absence of detectable cells (b) or plaques (e).
To test whether pre-existing CD44+Tet+CD4+ TM cells conferred protection, P + S K18-hACE2 mice, which express human angiotensin-converting enzyme 2 (ACE2) receptors mainly on epithelial cells, were injected i.n. and i.p. with PBS or a CD4-depleting antibody at days 0, 2, 4, 6, 9 and 12 after boosting i.n. with an unadjuvanted spike protein, followed by SARS-CoV-2 WA1/2020 virus infection at day 42 post-boosting i.n. Although i.n. boosted mice that were treated with PBS were protected against infection, as indicated by weight loss and survival, >50% of the mice treated with the CD4-depleting antibody succumbed at day 7 after viral infection (Fig. 5d) and had an increased infectious viral load in the lungs (Fig. 5e), indicating that loss of pre-existing CD4+ T cells eliminated the benefits of the intranasal booster. Thus, CD4+ T cells were required for the protective immunity conferred by the intranasal booster, at least in part by inducing respiratory IgA, IgG and tissue-resident CD8+ TM cell responses.
CD40 and TGFβ signaling drives lung IgA+ ASC differentiationWe next investigated the factors required for the antigen-specific IgA+ plasma cell development in the lower respiratory mucosa. To test whether CD40-mediated cognate interactions between CD4+ T cells and B cells in a TGFβ-rich niche were required for antigen-specific IgA+ ASC development, we injected P + S C57BL/6J mice with a CD40L-blocking antibody at days 0, 2, 4 and 6 post-boosting i.n. (Fig. 6a). Lung RBD-specific, CD38−GL7+ GC-like B cells, CD38+GL7−IgM−IgD− class-switched B cells and IgA+ ASCs were nearly absent (Fig. 6b) and BALF S1-specific IgA and IgG were substantially reduced (Fig. 6c), whereas serum S1-specific IgG remained unchanged (Fig. 6c) in CD40L antibody-treated mice compared with PBS-treated mice, indicating that CD40 signaling was required for the development of antigen-specific IgA+ plasma cells after an intranasal booster. TGFβ1 and TGFβ2 were increased, and lung CD19+ B cells expressed Aicda, which is required for CSR21,22,23,24,25, in P + S mice compared with intramuscular prime-only and i.n. boosted-only mice at days 18 and 21 post-priming i.m. (Fig. 6d–f). Intranasal and intraperitoneal treatment of P + S C57BL/6J mice with a TGFβ-neutralizing antibody at 14, 16, 18 and 20 d post-boosting i.n. notably reduced the RBD-specific IgA+ ASC numbers in the lungs (Fig. 6g,h), and specifically decreased the S1-specific IgA, but not S1-specific the IgG, levels in the BALF and serum compared with PBS-treated P + S mice (Fig. 6i). All isotype control antibodies corresponding to CXCR3, CXCL9, CXCL10, CD4, CD40L and TGFβ antibody depletion experiments did not change the number of CD44+Tet+CD4+ T cells, CD44+Tet+CD8+ T cells, RBD+CD38+GL7−IgM−IgD− class-switched B cells and RBD+IgA+ ASCs in the lung, and S1-specific IgA and IgG levels in the BALF and serum, compared with PBS treatment (Extended Data Fig. 8a–c), indicating that P + S mice treated with isotype control antibodies had comparable mucosal recall response to P + S mice treated with PBS. Thus, primed B cells developed into antigen-specific IgA+ ASCs with the help of CD40 and TGFβ stimulation on intranasal boosting with an unadjuvanted spike protein.
Fig. 6: CD40 and TGFβ signaling induce the development of mucosal IgA recall responses.
a, Schematic of the experimental setup showing C57BL/6J mice i.m. immunized with mRNA–LNPs at day 0, i.n. boosted with an unadjuvanted recombinant SARS-CoV-2 spike protein at day 14, followed by intraperitoneal treatment with PBS or a CD40L-blocking antibody at days 14, 16, 18 and 20 after intramuscular priming and lung, BALF and serum were analyzed at day 21 post-priming i.m. b, Number of extravascular IV−RBD+CD38−GL7+ GC-like B cells, IV−RBD+CD38+GL7−IgM−IgD− class-switched B cells and IV−RBD+IgA+ ASCs in the lungs of naive C57BL/6J mice (D0, n = 8), P + S mice treated with PBS (D21PBS, n = 10) or CD40L-blocking antibody (D21Ab, n = 10), analyzed by flow cytometry at day 21 post-priming i.m. as in a. c, ELISA measurements of SARS-CoV-2 S1-specific IgA and IgG in the BALF and serum from mice as in b. d, Schematic of the experimental setup showing C57BL/6 mice primed i.m. with mRNA–LNPs at day 0 that were boosted i.n. with spike protein at day 14, followed by lung and BALF analysis at days 16, 18 and 21 after priming i.m. e, ELISA measurement of TGFβ1 and TGFβ2 in BALF from naive mice (D0, n = 10); intramuscular prime-only (D16P, n = 9) and intranasal booster-only (D16S, n = 10) at day 16, and P + S mice at day 16 (D16P+S, n = 16), day 18 (D18P+S, n
Comments (0)