
Remember me
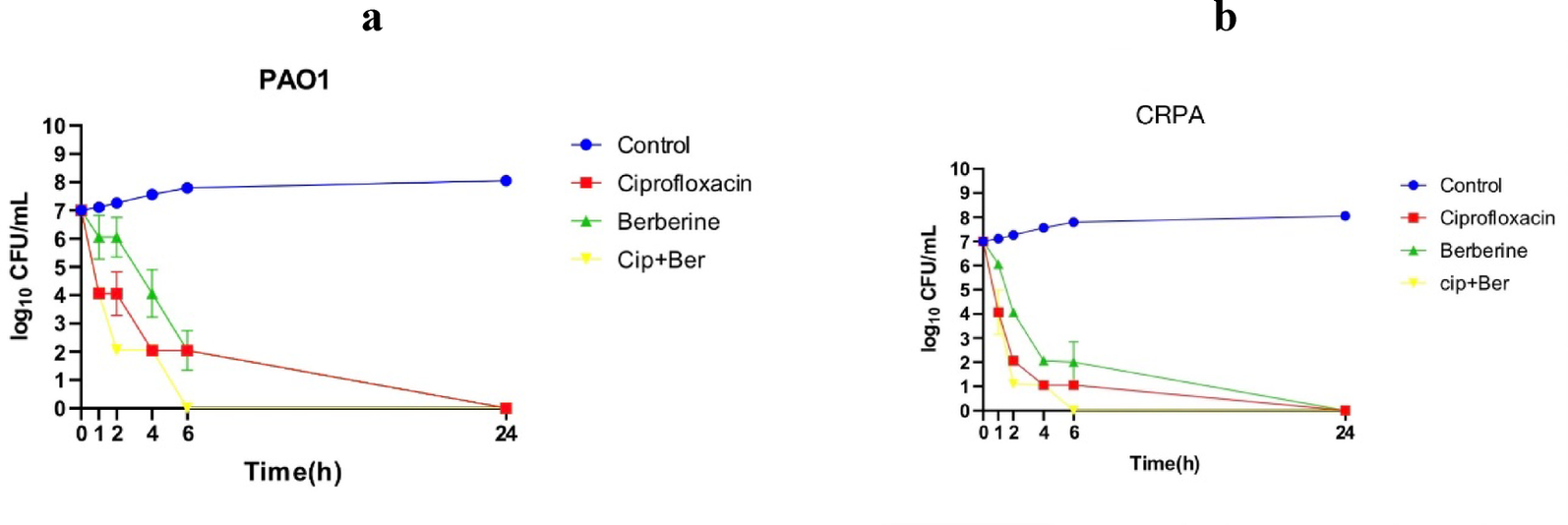





The cell shape of ZZQ-149 was short rod, and the width and length were in range of 0.78–1.85 μm and 1.38–6.04 μm, respectively (Fig. 1A; Table 1). The strain forms white colonies (Fig. 1B; Table 1), grows well in liquid LB medium with NaCl 1–8% (w/v) concentration but exhibits little mass growth in medium containing 10–30% NaCl (Fig S1). ZZQ-149 cells tend to aggregate and precipitate in 4–6% (w/v) NaCl. Studies indicate its viability in temperature between 16–47℃, with optimum growth between 28–37℃ (Fig S2). Furthermore, it can tolerate a pH range of 6.0–12.0, optimally thriving between pH 6.0–9.0 (Fig S3). ZZQ-149 forms PHA granules within cells following growth in halophilic medium with a 5% (w/v) NaCl concentration through an open, non-sterile fermentation process (Fig. 1).
Fig. 1
Cell and PHA granules under TEM observation and clone of ZZQ-149 on solid LB medium plate. (A) The dark circles with various shapes are cell of ZZQ-149 at different directions. The blight dots in the cells are PHA granules. (B) The clone of ZZQ-149 on the plate is white
Table 1 The differential characteristics of ZZQ-149T, and eight reference strains within the genus HalomonasZZQ-149’s grow occurs in ethanol, glycerol and D-mannitol as sole carbon source, yet it fails to thrive with D-galactose, D-xylose, and citrate as carbon source (Table 1 and Fig S4). It can grow with single nitrogen source including L-alanine, L-lysine, L-serine, L-valine and L-glycine, except for L-cysteine, L-glutamine and L-tryptophan (Table 1 and Fig S5). Growth in mediums with D-glucose and sucrose as sole carbon source leads to acid production; however, no acid production occurs with D-fructose as sole carbon source (Table 1 and Fig S4). Testing shows ZZQ-149’s ability to decompose gelatin, casein, starch and tween 80 (Table 1 and Fig S6).
In silico chemotaxonomic analysis reveals that ZZQ-149 harbors 249, 137, 60, 83, 267, 55, 54, 164, 23, 40 and 82 enzymes pertinent to carbohydrate metabolism, energy metabolism, lipid metabolism, nucleotide metabolism, amino acid metabolism, metabolism of other amino acids, glycan biosynthesis and metabolism, metabolism of cofactors and vitamins, metabolism of terpenoids and polyketides, biosynthesis of other secondary metabolites, and xenobiotics biodegradation and metabolism correspondingly (Fig. 2). Among the outlined eleven metabolic pathways, ZZQ-149 ranks eighth, fifth, fourth, seventh, second, fourth, seventh, sixth, sixth, fourth, and second among related species, respectively (Fig. 2). A detailed polar lipid analysis demonstrates that ZZQ-149 genome sequence encompasses the genes encoding for enzymes phosphatidyl glycero phosphate synthetase A (pgsA; E.C. 2.7.8.5) and phosphatidyl glycero phosphatase A (pgpA; E.C. 3.1.3.27) essential for the synthesis of phosphatidylglycerol (PG) from cytidine diphosphate (CDP)-diacyl glycerol (Fig S7A). The genome also contains the gene encoding for phosphatidylserine synthase (pssA; E.C. 2.7.8.8) that facilitates phosphatidylserine (PS) synthesis from CDP-diacyl-glycerol, while the synthesis of phosphatidyl-ethanolamine (PE) from PS is steered by phosphatidylserine decarboxylase (psd; E.C. 4.1.1.65) (Fig S7A). In addition, the genome incorporates the cardiolipin synthase gene (ClsA/B; E.C. 2.7.8.-) necessary for producing diphosphatidylglycerol (DPG; Cardiolipin) from PG, along with the gene of phospholipase A1 (E.C. 3.1.1.32) synthesizing 2-acylglycerophoserine from PS or the 2-acylglycerophoethanolamine from PE (Fig S7A). Furthermore, the gene of phospholipase A2 (E.C. 3.1.1.4), crucial for synthesizing 1-acylglycerophoethanolamine from PE, and the gene of 2-acylglycerophoethanolamine O-acyltransferase (E.C. 2.3.1.40), catalyzing the reaction process from 2-acylglycerophoethanolamine to PE are also present in the ZZQ-149 genome (Fig S7A). In comparison to ZZQ-149, the glycerophospholipid metabolisms are differently configured in four other strains as follows: (1) H. lutescens Q1UT harbors the cardiolipin synthase C gene (clsC; E.C. 2.7.8.-) (Fig S7B); (2) Although H. malpeensis YU-PRIM-29T possesses clsC, it lacks the 2-acylglycerophoethanolamine O-acyltransferase gene (E.C. 2.3.1.40) (Fig S7C); (3) Both H. nigrificans MBT G8648T and H. songnenensis NEAU-ST10-39T contain clsC gene but lack phospholipase A1/A2 gene (E.C. 3.1.1.32; E.C. 3.1.1.4) (Fig S7D and E); (4) H. piezotolerans NBT06E8T, H. rituensis TQ8ST, H. salicampi BH103T and H. zhuhanensis ZH2ST are devoid of the 2-acylglycerophoethanolamine O-acyltransferase gene (E.C. 2.3.1.40) (Fig S7F, G, H and I).
Fig. 2
Comparison of metabolisms among ZZQ-149 and its closest related species. The number of enzymes in eleven metabolism pathways of ZZQ-149 and other eight related species, including carbohydrate, energy, lipid, nucleotide, amino acid, glycan, cofactors and vitamins, terpenoids and polyketides, and xenobiotics
The distinctive attributes of ZZQ-149 differentiate it markedly from nearest related species. Initially, the white colonies of ZZQ-149 contrast with the yellow colonies of H. songnenensis NEAU-ST10-39T, H. malpeensis YU-PRIM-29T and H. piezotolerans NBT06E8T. Additionally, ZZQ-149 exhibits a larger cell size than H. salicampi BH103T and H. malpeensis YU-PRIM-29T. Its ability to grow across a broader salt concentration sets it apart from other species. ZZQ-149 also shares a similar temperature range to H. malpeensis YU-PRIM-29T, which is broader than that of other closely related species. However, it thrives in higher pH ranges compared to other species, except for H. salicampi BH103T. Unlike the eight closet species, ZZQ-149 can use different sole carbon and nitrogen sources, and demonstrate various enzymic activities alongside the capability to produce acid. ZZQ-149 possesses a unique metabolic pathway, with a particular emphasis on polar lipid metabolism. With a DNA G + C content of 52%, lower than other akin species, and the ability to synthesize PHA like H. lutescens Q1UT, this organism distinguishes itself further. Hence, the unique characteristics of ZZQ-149 assert its stand as a new Halomonas species.
ZZQ-149 genome featuresThe genomic sequence and characteristics of ZZQ-149 was derived using a synergy of PacBio and Illumina Hiseq2500 technologies. The PacBio’s whole-genome sequencing generated a total of 117,975 reads with a mean read length of 10,364 bp, facilitating a 317.44-fold genome coverage. Seamlessly, Illumina Hiseq2500’s genomic DNA sequencing provided a total of 7,443,122 reads, delivering an average read length of 150 bp and attaining 290-fold genome coverage. The composition of ZZQ-149 includes a circular chromosome (CP096973.1) and a plasmid pH149-1 (CP096974.1) (Fig. 3 and Table S1). The genome size of ZZQ-149 is smaller than the average Halomonas genus size of 4,003,450 bps, ranging between 1,136,546 bps at its minimum and 7,871,904 bps at its maximum. Of note, the concomitant range of gene number fluctuates between 1,137 and 7,872. Very few strains within the Halomonas genus contain plasmids, with ZZQ-149 being one of them, including the unnamed plasmid (CP080628.1) from H. qaidamensis XH36, plasmid pBAA-803-A (AP019515.1) from H. sulfidaeris ATCC BAA-803, and plasmid Pnrs2HaP11 (CP022738.1), Pnrs2HaP12 (CP022739.1) and Pnrs2HaP13 (CP022740.1) from H. ventosae NRS2HaP1. As an observation, pH149-1 (6, 107 bps) is considered a small plasmid. The genome annotation identifies that the pH149-1 plasmid encodes eight genes. These genes include a replication initiation protein (UYO76371.1), a helix-turn-helix domain-containing protein (UYO76372.1), an Eco29kI family restriction endonuclease (UYO76375.1), a DNA cytosine methyltransferase (UYO76376.1), a BrnT family toxin (M0220_17300), a BrnA antitoxin family protein (UYO76370.1), and two hypothetical protein (UYO76372.1 and UYO76373.1). Toxin-antitoxin systems are small loci found throughout bacteria and archaea and usually thought to mediate phage defence [16]. A putative toxin-antitoxin system, BrnT/ BrnA is only present in pH149-1 small plasmid, suggestive of possible novel biological functions and evolutionary adaptation within the Halomonas genus. Such genetic uniqueness is hypothesized to aid in the maintenance of plasmid residence within the host.
Fig. 3
Circle chromosome and plasmid genome of ZZQ-149. The circle genome of chromosome and plasmid were formed by Proksee [33] (https://proksee.ca). GC skew (innermost circle), GC content (second circle), and original features (i.e. CDS, tRNA, rRNA) extracted from a GenBank file (CP096973.1 and CP096974.1, third circle) are shown. Original features, such as CDS in outside of the backbone (gray) are from positive strand (+) of DNA, and the original features shown inside of backbone are from negative strand (-) of DNA. The putative toxin-antitoxin system, BrnT/BrnA is marked near corresponding genes of the plasmid
Phylogenetic position of ZZQ-149A 16 S rRNA gene sequence with 1,386 nucleotides was obtained from ZZQ-149 and was made publicly available on NCBI (https://www.ncbi.nlm.nih.gov/) under the accession number PP463991. Upon comparing the 16 S rRNA gene sequences of ZZQ-149 and other type strains, it was found that strain ZZQ-149 has the closest resemblance to H. zhaodongensis NEAU-ST10-25T, H. alkaliphila 18bAGT, H. andesensis LC6T and H. hydrothermalis Slthf2T with 99.19, 99.06, 98.84 and 98.82% sequence similarities, respectively. This comparison was made using the nucleotide similarity search program on the EzBioCloud server. Despite the similarities, the phylogenetic analysis which employed the NJ algorithm revealed that ZZQ-149 mostly similarly grouped with H. piezotolerans NBT06E8 within the expansive genus Halomonas cluster (Fig. 4). This cluster grouping was further supported through the phylogenetic trees based on the ML and MP algorithms, showing that ZZQ-149 fell within the genus Halomonas of the family Halomonadaceae (Fig S8). A separate phylogenetic tree based on single copy core genes from ninety-nine selected Halomonas strains (Table S2) also supported this claim, showing clustering of ZZQ-149 with H. nigrificans, H. zhaodongensis, H. andesensis, H. hydrothermalis, H. venusta, H. alkaliphile and H. humidisoli (Fig S9). The ANI values depicted a variance between ZZQ-149 and other species, with a range of 78.5%, seen with H. malpeensis (WHVL01.1), to a maximum of 87.1% with H. humidisoli (GCF_002286955.1) (Fig. 5 and Table S3). As the highest ANI between ZZQ-149 and other Halomonas species is less than 95%, it can be inferred that ZZQ-149 is a unqiue species which is present under the lowest standard for being classified as a new species. Based on its distinct physiological and biochemical characteristics, as well as its phylogenetic relationship and ANI with other species, ZZQ-149 has been confirmed as a novel species of Halomonas and has been named H. qinghailakensis. This is the first instance where a complete genome sequence of H. qinghailakensis has been obtained, exhibiting potential benefits for future phylogenetic and molecular research related to this new species.
Fig. 4
The phylogenetic tree constructed by neighbour-joining algorithm between strain ZZQ-149T and closely related taxa of genus Halomonas based on 16 S rRNA gene sequences. Red filled circles (●) indicate that the same nodes were also recovered by the maximum-likelihood and maximum-parsimony algorithms. Bootstrap values are shown on nodes as percentages of 1000 replicates for values over 70%. Pseudomonas aeruginosa DSMZ50071T (X06684) was used as an outgroup. The scale bar equals 0.02 changes per nucleotide position
Fig. 5
ANI between ZZQ-149 and other species of Halomonas genus. The ANI analysis was carried out by fastANI [22] which was integrated into PGCGAP [21]. The ANI value below 77% are shown with “NA”
PHA formation-related enzymes and pathways of H. qinghailakensis ZZQ-149The strain ZZQ-149 demonstrates potential for moderate PHA formation through open, non-sterile fermentation, highlighting its potential as a low-cost bacterial cellular factory to produce PHA. Our analysis of PHA formation-related enzymes and pathways involved performing a BLASTP search of all known enzymes using ZZQ-149’s entire protein sequence database.
This process yielded 59 candidate enzymes related to PHA formation, divided into 23 categories (Table S4). Considerably, 35 candidates corresponded to eight out of fourteen previously documented pathways [14], including pathway I, II, III, IV, VI, X, XI and XIII (Fig. 6 and Table S4). Conversely, the remaining 24 candidates were scattered across incomplete pathways including associated way, pathway V, VII, IX, and XII (Table S4). Interestingly, enzymes of pathway VIII and XIV were absent from ZZQ-149’s genome. Pathway I enzymes involve five PhaA (QBK40993.1) candidates including ZZQ-149_01220, ZZQ-149_00292, ZZQ-149_00272, ZZQ-149_02004 and ZZQ-149_01936, while pathway II comprises four FabG (TRM00464.1) homologs including ZZQ-149_03019, ZZQ-149_00686, ZZQ-149_02018 and ZZQ-149_01389. In comparison, eight AldD (S10772) candidates from pathway VII including ZZQ-149_03141, ZZQ-149_01758, ZZQ-149_02097, ZZQ-149_02265, ZZQ-149_00718, ZZQ-149_02421, ZZQ-149_02316 and ZZQ-149_02455 were discovered. Furthermore, ZZQ-149 hosts six ChnA (AAG10026.1) homologs consisting of ZZQ-149_01145, ZZQ-149_03246, ZZQ-149_00312, ZZQ-149_03167, ZZQ-149_01999 and ZZQ-149_00289, two ChnD (AAG10028.1) homologs consisting of ZZQ-149_02266 and ZZQ-149_02491, and four ChnE (AAG10022.1) homologs consisting of ZZQ-149_02102, ZZQ-149_03175, ZZQ-149_02287 and ZZQ-149_02418 from pathway IX. Three FadA (QVM89852.1) candidates from pathway X (ZZQ-149_00263, ZZQ-149_01273 and ZZQ-149_00264) and three LeuB (UNS57662.1) candidates from pathway XII (ZZQ-149_00048, ZZQ-149_02302 and ZZQ-149_02541) were also identified in ZZQ-149’s genome. Proteins, including ZZQ-149_00448, ZZQ-149_03018, ZZQ-149_01517, ZZQ-149_01125, ZZQ-149_02031, ZZQ-149_00264, and ZZQ-149_01666 were identified as the single homolog copy of PhaC (AEI76811.1) of pathway I, FadD (ARU38287.1) of pathway III, IlvA (NP_418220.1) and ThrC (CAD6022432.1) of pathway VI, DhaT (CAC9684055.1) of pathway VII, FadB (QVM89853.1) of pathway X, IdhA (EDV68311.1) of pathway XI, LeuC (UNS57713.1) and LeuD (UML70839.1) of pathway XII, respectively.
Fig. 6
PHA-synthesis pathways in ZZQ-149. PHA formation-related enmzyme candidates and potential corresponding pathways are speculated consulting the pathway in literature [14] and listed in Table S4. The serial numbers stand for the candidates of published enzymes in the above literature, and the homologs are used to speculate respective pathway of PHA synthesis in ZZQ-149
To analyze the evolutionary relationship and functions of enzymes, we have constructed phylogenetic trees from their nucleotide sequences and unnearthed functional domains using their protein sequences. Enzymes with two or more candidates underwent this analysis. Two PhaA (QBK40993.1) candidates, ZZQ-149_02004 and ZZQ-149_01936, both beta-ketoadipyl CoA thiolase, contain PRK09050 domain cluster alongside the beta-ketothiolase BktB candidate, ZZQ-149_00272, which inhabits the PRK09051 superfamily domain in cluster I (Fig S10A). Cluster II houses PhaA and ZZQ-149_01220 (putative acyltransferase) with a shared PRK05790 domain, and includes ZZQ-149_00292, an acetyl-CoA C-acetyltransferase with a PRK06954 superfamily domain (Fig S10A). Phylogenetic tree of FabG (TRM00464.1) and its candidates splits into three branches. Cluster I include ZZQ-149_03019 (3-ketoacyl-(acyl-carrier-protein) reductase with a fabG domain), ZZQ-149_02018 (benzoate diol dehydrogenase with BenD superfamily domain) and FabG (3-oxoacyl-ACP reductase with fabG domain). Cluster II and III include ZZQ-149_00686 (NADP-dependent 3-hydroxy acid dehydrogenase with YdfG domain) and ZZQ-149_01389 (Rossmann-fold NAD(P)(+)-binding protein with NADB_Rossmann superfamily domain), respectively (Fig S10B). ChnA (AAG10026.1), ZZQ-149_00312 (NAD(P)-dependent dehydrogenase), ZZQ-149_01999 (NAD(P)-dependent dehydrogenase) and ZZQ-149_03167 (3-ketoacyl-(acyl-carrier-protein) reductase) share the same FabG domain. However, ChnA and ZZQ-149_00312 gather with ZZQ-149_03246 (3-hydroxybutyrate dehydrogenase with PHB_DH superfamily domain) in cluster I, ZZQ-149_01999 and ZZQ-149_03167 get together with ZZQ-149_00289 (Rossmann-fold NAD(P)(+)-binding protein with NADB_Rossmann superfamily domain) in cluster III, leaving ZZQ-149_01145 (acetoacetyl-CoA reductase with AcAcCoA_reduct superfamily domain) alone in cluster II (Fig S10C). In case of ChnD (AAG10028.1) and ZZQ-149_02491 (cinnamyl alcohol dehydrogenase with CAD1 domain), they clump together in cluster I, and with ZZQ-149_02266 (cinnamyl alcohol dehydrogenase with CAD3 domain) in cluster II (Fig S10D). ZZQ-149_03175, ZZQ-149_02287, ZZQ-149_02102 and ChnE (AAG10022.1) are mitochondrial succinate-semialdehyde dehydrogenase with an ALDH_F5_SSADH_GabD domain. However, only ZZQ-149_03175, ZZQ-149_02287 and ZZQ-149_02102 cluster together in cluster I, while ChnE links with ZZQ-149_02418 (succinate-semialdehyde dehydrogenase with an ALDH_SSADH1_GabD1 domain) in cluster II (Fig S10E). AldD (S10772), ZZQ-149_03141 (betaine aldehyde dehydrogenase with a PRK13252 domain), ZZQ-149_02097 (NADP+-dependent gamma-glutamyl-gamma-aminobutyraldehyde dehydrogenase with an ALDH_GABALDH-PuuC domain), ZZQ-149_02265 (NAD+-dependent acetaldehyde dehydrogenase with an ALDH_ACDHII_AcoD-like domain) and ZZQ-149_01758 (NAD(P)+-dependent aldehyde dehydrogenase with an ALDH-SF superfamily domain) group together in cluster (I) Yet, ZZQ-149_02316, another ALDH-SF superfamily domain-containing candidate is the sole inhabitant of cluster (II) ZZQ-149_02421 (methylmalonate semialdehyde dehydrogenase with an ALDH_F6_MMSDH domain), ZZQ-149_00718 (NADP+-dependent alpha-ketoglutaric semialdehyde dehydrogenase with an ALDH_KGSADH-YcbD domain) and ZZQ-149_02455 (succinylglutamic semialdehyde dehydrogenase with an astD domain) assemble in cluster III (Fig S10F). FadA (QVM89852.1) share a cluster with its homologs, ZZQ-149_00263 (3-ketoacyl-CoA thiolase with a fadA superfamily domain) and ZZQ-149_01273 (acetyl-CoA C-acetyltransferase with a PRK06445 superfamily domain) (Fig S10G). In cluster I, LeuB (UNS57662.1) and ZZQ-149_00048, both 3-isopropylmalate dehydrogenase with a PRK00772 domain, group together. Stimultaneously, ZZQ-149_02302 (isocitrate dehydrogenase with a PRK08997 domain) and ZZQ-149_02541 (isocitrate/isopropylmalate dehydrogenase with an iso_dh superfamily domain) cluster gather in cluster II (Fig S10H). Our analysis of the evolutionary relationships and functional domains reveals that a single PHA-formation enzyme can have more than two homologous proteins each with different domains. The divergent roles of these candidate proteins within the PHA synthetic process likely derive from their individual functions.
The cooccurence of several pathways suggests that ZZQ-149 could serve as an optimal chassis cell for PHA production. Nonetheless, an in-deep exploration of certain areas could enhance the quality and yield of PHA. These include actual the roles of enzymes, mechanism of coexisting, distinct pathways as demonstrated in Fig. 6, pathway incompleteness, the simultaneous presence of various enzyme candidates with similar functionalities as seen in Fig S10. Furthermore, certain enzyme candidates, categorized into two specified pathways based on query enzyme sequences in BLASTp analysis and annotated functions, such as enzymes 1, 3 and 14 might be pivotal in PHA formation, as detailed in Table S4. Enzyme candidates with an identity of less than 30% or coverage below 60%, or annotated as hypothetical proteins, could be PHA- specific enzymes or may not participate in PHA synthesis in ZZQ-149 (Table S4).
Among ZZQ-149 and its closest related species, PHA production has only been reported in ZZQ-149 and H. lutescens Q1UT (Table 1). An examination of PHA formation-related enzymes reveals that Q1UT lacks one homolog (ZZQ-149_02018) of FabG (TRM00464.1) from pathway II, the only homolog (ZZQ-149_02031) of DhaT (CAC9684055.1) from pathway VII, the candidate (ZZQ-149_03160) of PhaJ (QGH90118.1) from pathway II and the candidate (ZZQ-149_02851) of ChnC (AAG10029.1) from pathway IX, as shown in Table S5. This disparity among PHA-synthesizing enzymes might lead to various types of PHA granule formation in ZZQ-149 and Q1UT.
Comments (0)