
Remember me



Our study yielded several salient findings. First, we report that NMR brain tissue is a net scavenger of ROS when mitochondria are respiring on all combinations of substrates tested, including states II and III respiration through complex I or complex II of the mitochondrial electron transport system (ETS). Second, the bulk of this scavenging is attributable to the thioredoxin-scavenging pathway, as an antagonist of this enzyme reduces much of the net scavenging capacity of NMR brain. Third, the capacity of NMR brain to scavenge ROS is Ca2+ dependent and decreases with increasing Ca2+.
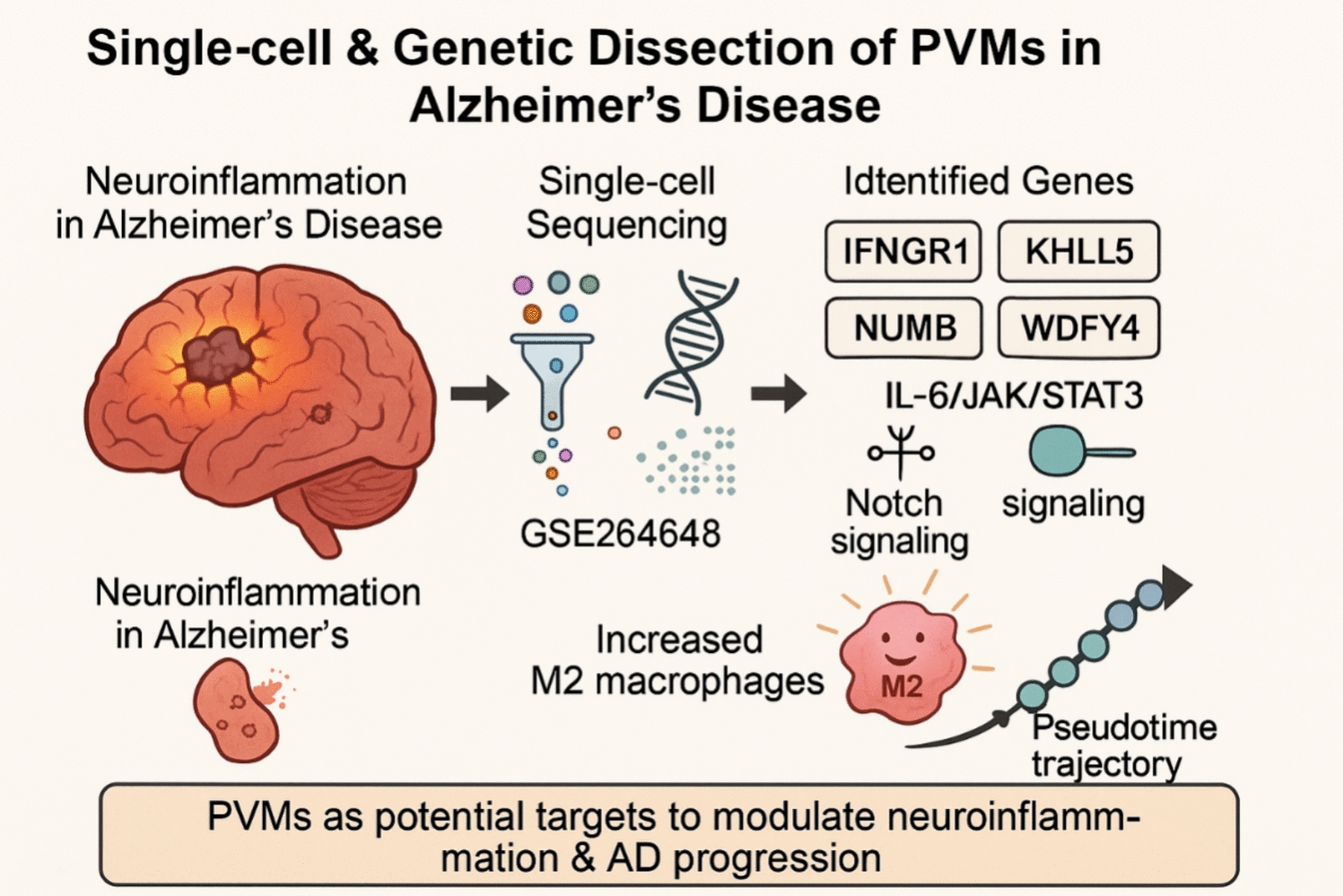
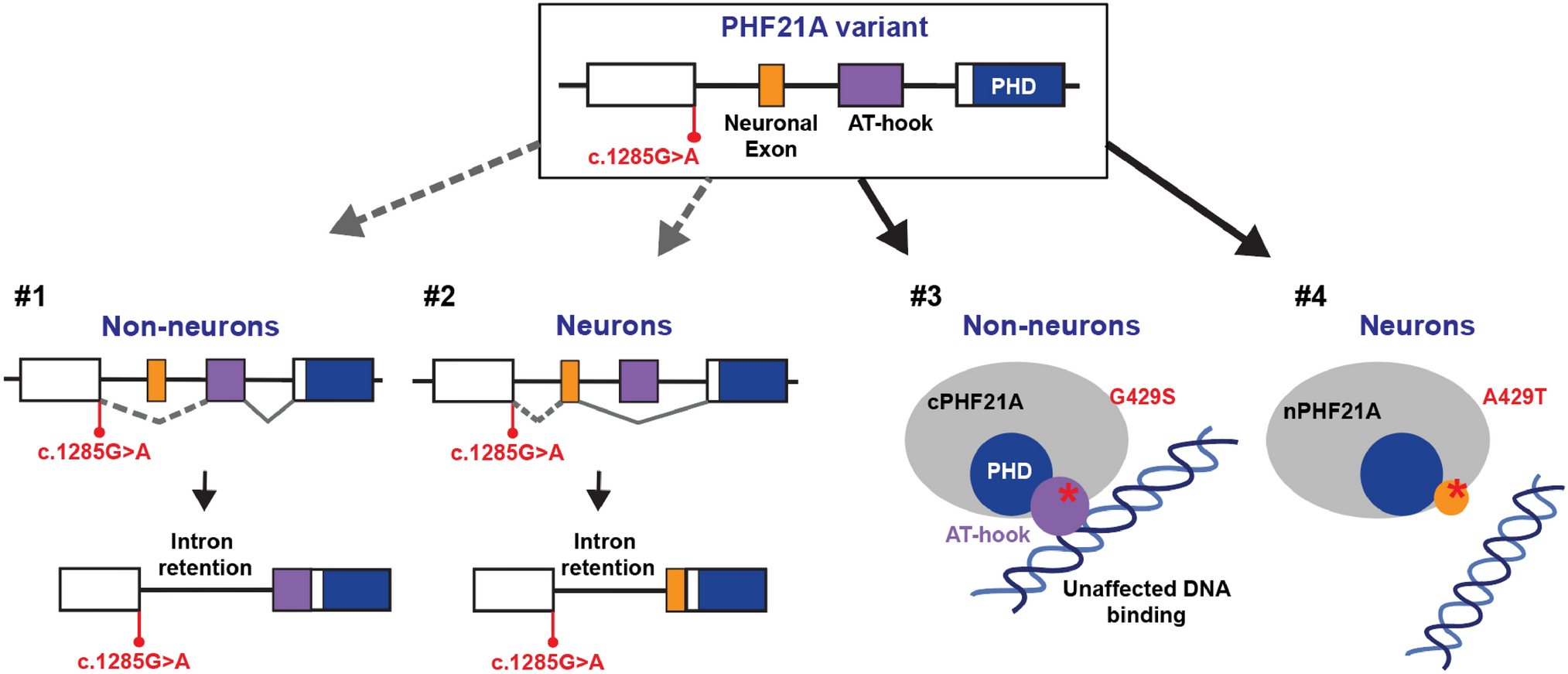
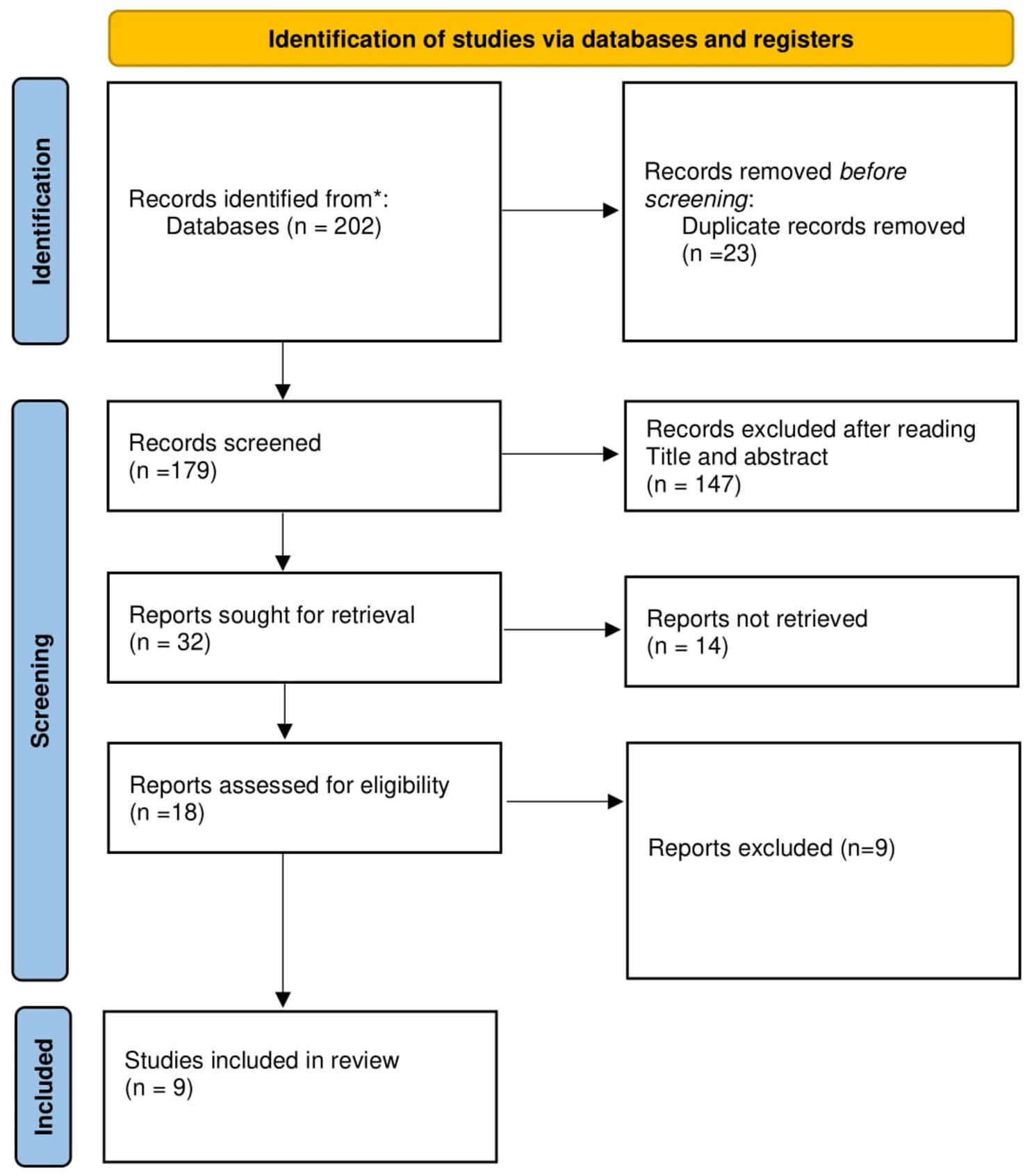
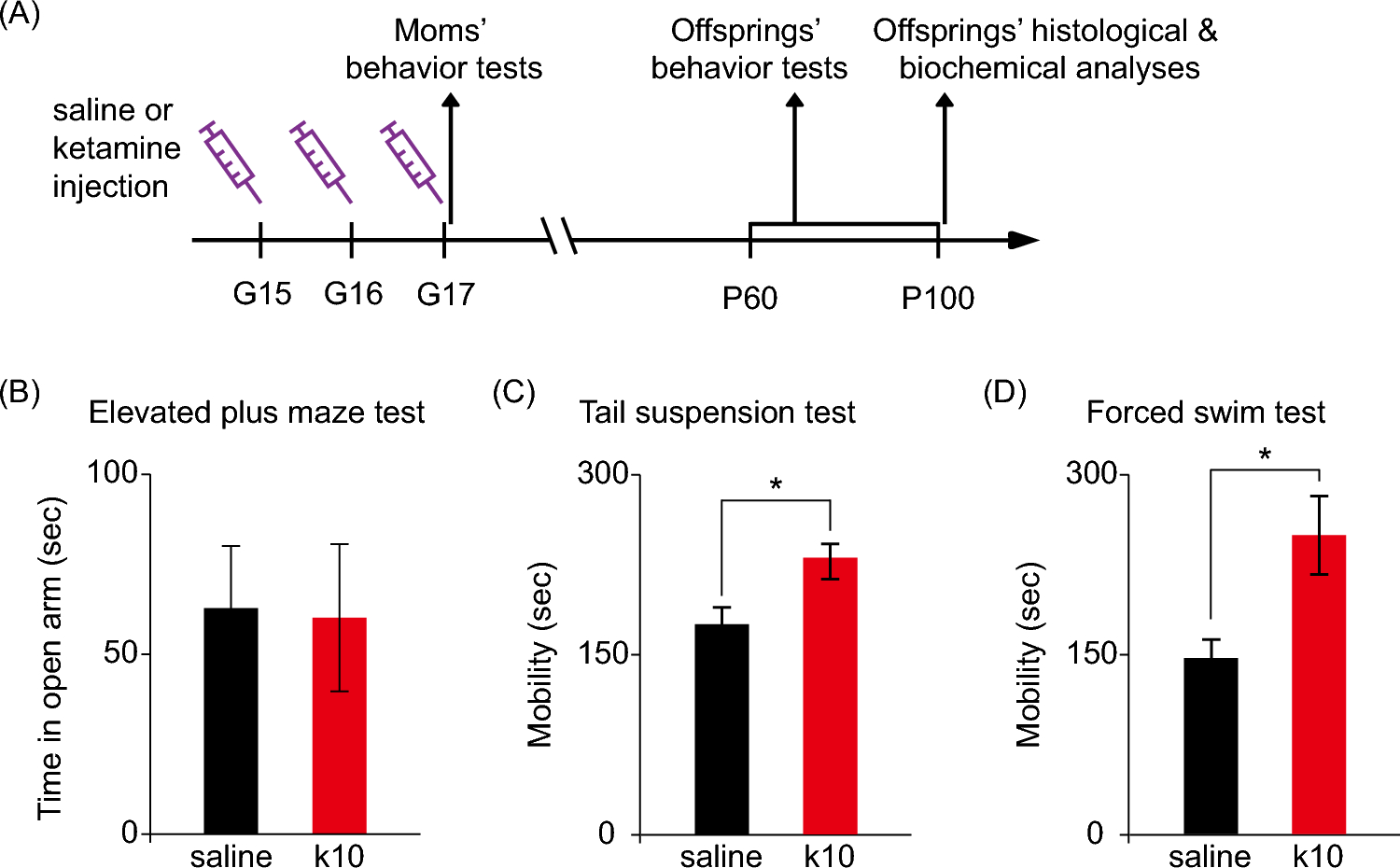
NMR Brain Tissue Is a Net Consumer of ROSWhen respiring on substrates for either complex I (Fig. 1; F(3,48) = 65.90, p < 0.0001; n = 7 each) or complex II (Fig. 2; F(3,24) = 33.76, p < 0.0001; n = 7 each) of the mitochondrial ETS, NMR brain mitochondria are net consumers of H2O2. The highest rate of consumption is observed in complex I-fuelled mitochondria respiring in state III (Fig. 1; GMD). In most cases, consumption is higher in conditions where mitochondrial respiration is highest, i.e., when mitochondria are energized with ADP to spark state III respiration. However, ADP addition does not change H2O2 consumption relative to succinate-induced leak respiration (Fig. 2; F1, 44 = 0.0048, p = 0.9447), unlike for complex I-fueled respiration (Fig. 1; F1, 46 = 87.96, p < 0.0001). Rotenone (which blocks reverse electron transport from complex II to I) increases H2O2 consumption by 200% during succinate-fuelled respiration (F1, 46 = 24.86, p = 0.0012).
Fig. 1
Summary of the impact of increasing Ca2+ on cellular scavenging of exogenous H2O2 by naked mole-rat cortical homogenates respiring on substrates for complex I of the mitochondrial electron transport system. Homogenates were exposed to 50 µl of 10-µm H2O2 and provided with glutamate (G) and malate (M), ± ADP (D), ± auranofin (AU), an inhibitor of thioredoxin reductase. Data are mean ± SEM from 7 biological replicates. Significance was determined using a one-way ANOVA and Tukey’s test (p < 0.05)
Fig. 2
Summary of the impact of increasing Ca2+ on cellular scavenging of exogenous H2O2 by naked mole-rat brain cell homogenates respiring on substrates for complex II of the mitochondrial electron transport system. Homogenates were exposed to 50 µl of 10-µm H2O2 and provided with succinate (S) ± ADP (D) ± rotenone (R), a specific inhibitor of mitochondrial complex I. Data are mean ± SEM from 7 biological replicates. Significance was determined using a one-way ANOVA and Tukey’s test (p < 0.05)
It is important to note that hypoxia-intolerant mammal brain is also a net consumer of ROS, with net consumption of exogenous H2O2 by isolated rat and mouse brain mitochondria occurring at ~ 1–12 nmol/mg/min, depending on the substrates utilized (Zoccarato et al. 2004; Drechsel and Patel 2010; Starkov et al. 2014). However, when converted to common units, our reported rate of ROS scavenging by NMR brain is ~ four-fold higher than in these hypoxia-intolerant mammals. Importantly, our study utilized homogenized cortical tissue, whereas these previous studies used isolated brain mitochondria. This is an important consideration because NADPH, which is a key substrate for both the glutathione- and thioredoxin-scavenging systems (Eaton and Pamenter 2022a), may be lost in the preparation of isolated mitochondria. It is also important to note that cytosolic ROS scavengers such as catalase and vitamins C and E may play an important role in scavenging ROS, and this contribution would be lost in experiments using isolated mitochondria. Further experiments are warranted to test the involvement of these systems in NMR brain ROS scavenging and to compare ROS scavenging rates between species using a common tissue preparation.
Nonetheless, this apparently high capacity to scavenge ROS in NMR brain is like our previous reports in NMR skeletal and cardiac muscle (Munro et al. 2019). Interestingly, a similarly high capacity to detoxify ROS has been reported in various tissues of another long-lived and hypoxia-tolerant animal. For example, the arctic clam (Arctica Islandica) has considerably greater ROS-scavenging capacity than shorter-lived hypoxia-intolerant clam species (Munro et al. 2023). Additional studies on tissues are warranted to determine if this is a common capacity in other hypoxia-tolerant species.
NMR Brain ROS Scavenging Is Largely Mediated by ThioredoxinOur results suggest a major role for the thioredoxin-scavenging pathway in NMR brain. Specifically, we evaluated the effect of treating cellular homogenates respiring on GMD with the thioredoxin reductase inhibitor auranofin (Saccoccia et al. 2014) using a two-way ANOVA and found that auranofin reduces the net scavenging capacity of H2O2 by NMR brain by > 80% (Fig. 1; F1, 6 = 262.8, p < 0.0001). However, further reductions in ROS-scavenging capacity are observed with increasing Ca2+ addition (see below), suggesting that additional scavenging pathways, which are Ca2+ sensitive, remain active. Indeed, if all scavenging capacity were abolished by auranofin we would predict a net increase in ROS generation from a preparation of actively respiring mitochondria treated with 50 µl of 10-µm H2O2. It is likely that glutathione peroxidase plays an important role here, as we have previously reported changes in glutathione metabolism in hypoxic NMR brain (Cheng et al. 2022), and because this system is specifically inhibited by Ca2+ (see below), which does not inhibit the thioredoxin system (Starkov et al. 2014). In addition, changes in ROS generation likely make a small contribution to the changes we report as ROS generation is inherently linked to cellular O2 consumption (Eaton and Pamenter 2022b; Cheng and Pamenter 2021) and because Ca2+ can inhibit NMR brain mitochondrial respiration under certain substrate conditions (Cheng et al. 2024). However, the rate of natural ROS generation from NMR cortical brain homogenates is ~ 1/100th of the rate of consumption observed in the presence of the large bolus of H2O2 provided in the present study. Therefore, it is likely that the large majority of the change in ROS consumption induced by Ca2+ addition is due to changes in ROS-scavenging systems.
Across mammals in general, there is debate in the literature about the primary mediators of ROS scavenging in brain. One study reported that the glutathione system is a major component of ROS scavenging in isolated rat brain mitochondria (Zoccarato et al. 2004), but that a significant additional scavenging capacity (~ 40%) is inhibited by the peroxidase inhibitor arsenite, which does not target the glutathione pathway. Conversely, another study in isolated rat brain mitochondria using a different experimental approach found that ~ 25% of ROS scavenging capacity was attributable to the glutathione pathway (and a similar % was blocked by arsenite), with the remainder attributable to the thioredoxin pathway (Drechsel and Patel 2010). A third study, conducted in isolated mouse brain mitochondria, largely agrees with the 2nd rat study and supports a primary role for thioredoxin in detoxifying brain mitochondria (Starkov et al. 2014), in good agreement with our results.
Ca2+ Impairs ROS Scavenging in NMR BrainLastly, we observed that Ca2+ inhibits H2O2 consumption in permeabilized NMR brain respiring on all substrates except succinate-induced leak respiration (Figs. 1 and 2; F3, 114 = 92.41, p < 0.0001). In addition, H2O2 consumption is more sensitive to Ca2+ in mitochondria fueled through the complex II (succinate/rotenone) pathway than the complex I (glutamate/malate) pathway. These results are generally consistent with experiments in isolated brain mitochondria from hypoxia-intolerant mammals. For example, in isolated rat cortical mitochondria, ROS scavenging is 80% lower in preparations with EGTA (a Ca2+ chelator) in the respiration medium, or when the mitochondrial Ca2+ uniporter (which mediates mitochondrial Ca2+ uptake) is blocked (Zoccarato et al. 2004). This reduction is mediated in part through direct suppression of the glutathione reductase pathway by Ca2+ (Zoccarato et al. 2004). Similar results have been reported in other studies, with one study reporting an EC50 for Ca2+ inhibition occurring at 42 µM Ca2+ (Drechsel and Patel 2010). Our results suggest that NMR brain can accumulate a somewhat higher level of Ca2+ before reaching the 50% inhibition threshold (Cheng et al. 2024). This ability may reflect the enhanced Ca2+ buffering capacity of relatively larger brain mitochondria in this species, which have a larger mitochondrial area and volume, higher cristae density, and more cristae junctions per mitochondria and per unit area than in mice (Cheng et al. 2024).
Comments (0)