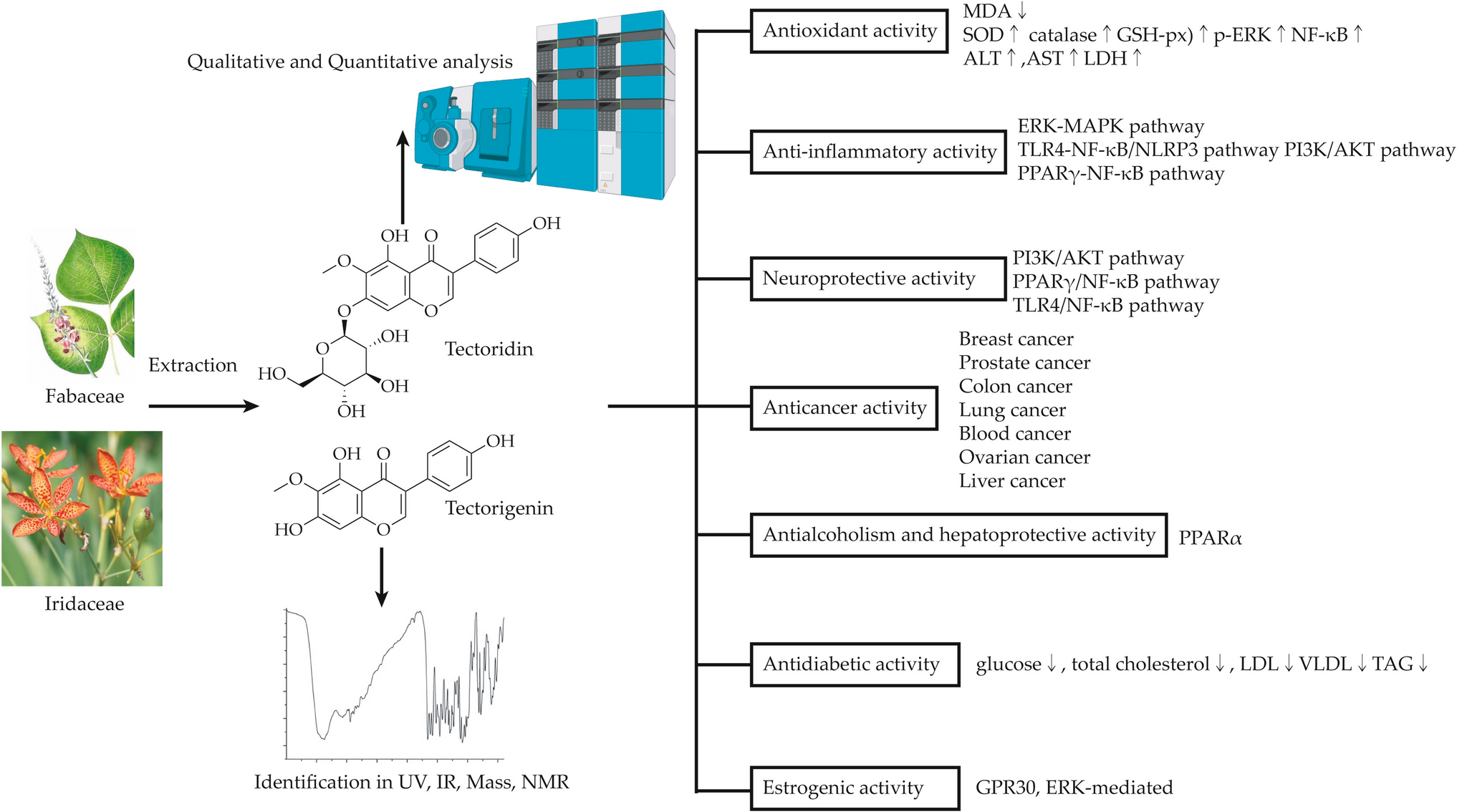
Antioxidant Activity
Tectoridin and tectorigenin have demonstrated efficacy in ameliorating carbon tetrachloride (CCl4)-provoked hepatotoxicity in rodent models, as evidenced by a pronounced suppression of serum transaminase elevation and reduced malondialdehyde (MDA) levels quantified via the thiobarbituric acid assay (Jung et al. 2004). Furthermore, both compounds markedly elevated the activity of hepatic antioxidant defense systems, including cytosolic superoxide dismutase (SOD), catalase, and glutathione peroxidase (GSH-Px), in CCl4-intoxicated rats (Jung et al. 2004). This antioxidant potentiation was similarly observed in Chinese hamster lung fibroblast (V79-4) cell line, corroborating the cytoprotective mechanism across experimental models (Kang et al. 2005; Zhang et al. 2016).
Lee et al. (2003) reported that intraperitoneal administration of tectorigenin (100 mg/kg) markedly suppressed the elevation of plasma alanine aminotransferase (ALT), aspartate aminotransferase (AST), and lactate dehydrogenase (LDH) activities in murine models of carbon tetrachloride-induced hepatic injury. Oral administration of tectoridin not only attenuated β-glucuronidase activity but also enhanced hepatic glutathione (GSH) levels and glutathione S-transferase (GST) activity in CCl4-exposed mice (Lee et al. 2003). Furthermore, tectorigenin mitigated hydrogen peroxide (H2O2)-triggered apoptotic cell death and G2/M-phase cell cycle arrest in V79-4 fibroblasts, a cytoprotective effect attributed to its antioxidant properties and activation of the extracellular signal-regulated kinase (ERK) signaling pathway. These findings suggest that tectorigenin and tectoridin possess antioxidative properties in cells and hepatoprotective activities in CCl4-intoxicated rats (Jung et al. 2004).
Han et al. (2012) performed in vitro assays to assess the antioxidant potential of tectoridin and tectorigenin, quantifying their reducing capacity alongside radical scavenging efficacy against superoxide anions, hydroxyl radicals, and 1,1-diphenyl-2-picrylhydrazyl (DPPH), as well as their inhibitory effects on lipid peroxidation-benchmarked against ascorbic acid and butylated hydroxytoluene. Lee et al. (2000) demonstrated that tectorigenin’s in vitro antioxidant properties, demonstrating its capacity to neutralize DPPH radicals, quench superoxide anions generated via the xanthine/xanthine oxidase system, and suppress lipid peroxidation in rodent microsomal preparations through enzymatic and non-enzymatic mechanisms. Zhang et al. (2016) further established tectorigenin’s in vivo antioxidant efficacy, showing attenuation of hydrogen peroxide-induced cytotoxicity in V79-4 cells alongside enhanced catalase activity and upregulated protein expression. Notably, tectorigenin’s cytoprotective action against oxidative stress was diminished upon pharmacological inhibition of catalase activity. Mechanistic investigations revealed tectorigenin’s ability to potentiate ERK phosphorylation and nuclear translocation of nuclear factor kappa-B (NF-κB), whereas pharmacological blockade of these pathways compromised its antioxidative efficacy.
Han et al. (2012) further suggested that the antioxidant activity in all experimental systems followed this order: tectorigenin sodium sulfonate > tectorigenin > tectoridin. tectorigenin also protected the Vero cell line (normal monkey kidney) from hydrogen peroxide-induced injury. The antioxidant properties of tectorigenin may help mitigate streptozotocin-induced toxicity and contribute to its hypoglycemic and hypolipidemic effects (Lee et al. 2000).
Tectorigenin and tectoridin demonstrate remarkable antioxidative and hepatoprotective properties, significantly mitigating CCl4-induced hepatic damage by reducing oxidative stress and enhancing key antioxidant enzymes such as SOD, catalase, and GSH-px. Their efficacy extends to the inhibition of apoptotic cell formation, lipid peroxidation, and cell cycle arrest, while promoting pathways like ERK and NF-κB for cellular protection. These findings underscore their therapeutic potential in oxidative stress-related conditions, liver injuries, and metabolic disorders, marking them as promising candidates for further pharmacological exploration.
Anti-inflammatory Activity
Numerous studies have shown that tectoridin and tectorigenin possess significant anti-inflammatory activity through the regulation of various pathways, including the ERK-mitogen-activated protein kinase (MAPK) pathway (Dong et al. 2023), Toll-like receptor 4 (TLR4)-NF-κB/nucleotide oligomerization domain-like receptor protein-3 (NLRP3) pathway (Niu et al. 2022), phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) pathway, and peroxisome proliferator-activated receptor gamma (PPARγ)-NF-κB pathway (Yao et al. 2021).
Dong et al. (2023) demonstrated that tectoridin ameliorated ankle joint pathology and synovial tissue degeneration in murine models of complete Freund’s adjuvant (CFA)-provoked arthritis. Furthermore, tectoridin attenuated the release of pro-inflammatory cytokines in both tumor necrosis factor-α (TNF-α) stimulated human fibroblast-like synoviocytes (HFLS) in vitro and CFA-challenged arthritic mice in vivo. Concurrently, tectoridin exhibited marked suppression of aberrant ERK and c-Jun N-terminal kinase (JNK) phosphorylation across experimental models, underscoring its anti-arthritic efficacy through inhibition of MAPK-dependent inflammatory signaling pathway.
Both tectoridin and tectorigenin have been shown to inhibit prostaglandin E2 production and cyclooxygenase-2 (COX-2) induction in rat peritoneal macrophages, indicating their anti-inflammatory activities by suppressing these inflammatory mediators (Kim et al. 1999; Pan et al. 2008). Tectorigenin, in particular, exhibits more potent inhibition of prostaglandin E2 production compared to tectoridin (Kim et al. 1999). Furthermore, the anti-inflammatory potential of tectoridin and tectorigenin has been investigated in macrophages exposed to interferon-γ and lipopolysaccharide (IFN-γ/LPS) co-stimulation (Pan et al. 2008). In IFN-γ/LPS-activated RAW 264.7 murine macrophages, tectoridin demonstrated dose-dependent suppression of inducible nitric oxide synthase (iNOS) expression, concomitant with attenuated nitric oxide (NO) synthesis and diminished interleukin-1β (IL-1β) release (Pan et al. 2008).
Tectorigenin’s anti-inflammatory effects are attributed to its ability to block the activation of NF-κB (Pan et al. 2008). Moreover, tectoridin has shown an inhibition rate of 53.71% at a concentration of 10 μM in anti-inflammatory study (Liu et al. 2022). Research indicates that tectoridin reduces LPS-induced increases in NO, interleukin-6 (IL-6), and interleukin-18 (IL-18) in macrophages, as well as decreases TNF-α, IL-6, and IL-1β levels in serum (Niu et al. 2022). Histopathological study have also highlighted tectoridin’s hepatoprotective and lung-protective effects (Niu et al. 2022).
Emerging evidence posits tectorigenin as a prospective therapeutic agent for stroke mitigation, although its precise mechanisms remain incompletely characterized. In a mechanistic investigation, tectoridin was evaluated for its capacity to counteract inflammation, oxidative stress, and apoptosis in HT-22 neuronal cells subjected to oxygen–glucose deprivation/reperfusion (OGD/R) injury. tectoridin treatment robustly enhanced cellular viability, attenuated apoptotic cell death, and suppressed ROS generation alongside IL-1β, IL-6, and TNF-α production under OGD/R conditions. Concurrently, tectoridin upregulated PI3K/AKT signaling, elevated PPARγ expression, and downregulated NF-κB pathway activation in this cellular model (Yao et al. 2021).
Both tectoridin and tectorigenin exhibit low cytotoxic effects. In a macrophage cell line (RAW 264.7) activated with interferon-γ and LPS, pretreatment with tectorigenin inhibited iNOS and NO production, as well as IL-1β secretion, in a dose-dependent manner, without affecting TNF-α secretion. Tectorigenin also dose-dependently decreased COX-2 and prostaglandin E2 production. Compared to tectoridin, tectorigenin demonstrated greater inhibition of NO production and inflammatory responses in activated monocyte/macrophage cells.
Research has demonstrated that tectorigenin significantly reduces neutrophil migration to lung tissue following LPS exposure, as indicated by a notable decrease in myeloperoxidase activity. Furthermore, eosinophilic inflammation, a hallmark of allergic asthma, was effectively mitigated by tectorigenin in an asthma model (Ma et al. 2014). A study shown significantly lowered eosinophil infiltration around bronchial and vascular regions in asthmatic mice, highlighting its potential as a therapeutic agent for managing eosinophilic inflammation in asthma and related conditions (Guo et al. 2024).
Tectoridin and tectorigenin exhibit significant anti-inflammatory activity by modulating key signaling pathways, such as ERK-MAPK, TLR4-NF-κB/NLRP3, PI3K/AKT, and PPARγ-NF-κB. These compounds suppress pro-inflammatory mediators, including COX-2, prostaglandin E2, and NO, and reduce cytokine secretion, such as TNF-α, IL-6, and IL-1β. Their effects extend to protective actions against oxidative and inflammatory damage in diverse models, including arthritis, macrophage activation, and stroke-related injuries. Furthermore, their low cytotoxicity and robust anti-inflammatory properties underscore their potential as promising therapeutic agents for managing inflammatory diseases.
Neuroprotective Activity
Research has established that tectoridin and tectorigenin mediate neuroprotective effects, primarily by orchestrating the activity of pivotal regulatory proteins—including phosphorylated protein kinase B (p-AKT), B-cell lymphoma 2 (Bcl-2)/Bcl-2-associated X protein (Bax), hypoxia-inducible factor-1α (HIF-1α), and phosphorylated mammalian target of rapamycin (p-mTOR)—through the activation of PI3K/AKT and PPARγ/NF-κB signaling pathways (Noh et al. 2019).
Chen et al. (2023) conducted in-depth investigations showing that tectoridin effectively reduces the overproduction of reactive oxygen species (ROS) and intracellular Ca2+ levels while enhancing mitochondrial membrane potential (MMP). These actions help mitigate ROS-mediated mitochondrial dysfunction. Additionally, tectoridin inhibits apoptosis by upregulating the expressions of p-AKT, Bcl-2/Bax, p-mTOR, and nuclear factor erythroid 2-related factor 2 (Nrf2). Tectoridin also reduces the expressions of Bcl-2/Bax, p-AKT, p-mTOR, Nrf2, heme oxygenase-1 (HO-1), nicotinamide adenine dinucleotide (phosphate) reduced:quinone oxidoreductase (NQO1), and glutamate-cysteine ligase (GCLM), thereby protecting rat pheochromocytoma cells (PC12 cells) from oxygen–glucose deprivation/reoxygenation-induced damage through the activation of PI3K/AKT signaling pathways.
Tectorigenin plays a significant role in neuroprotection by inducing the transcription of HIF-1α and preventing its degradation, which is crucial for regulating erythropoietin during neuronal treatment (Gong et al. 2017). Furthermore, tectorigenin inhibits mitochondrial processing peptidase (MPP)-induced alterations in Bax and Bcl-2 levels and increases ROS levels, along with the expression of nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) oxidase, superoxide dismutase, catalase, and glutathione peroxidase in human neuroblastoma cell line (SH-SY5Y) (Gong et al. 2017).
Both tectoridin and tectorigenin have demonstrated neuroprotective effects on dopaminergic neurons in vitro (SH-SY5Y cells) and in vivo Parkinson’s disease (PD) models. Gong et al. (2017) found that tectorigenin provided neuroprotection against PD in MPP-treated SH-SY5Y cells, while Lim et al. (2018) suggested that tectoridin may suppress neuronal inflammation by downregulating inflammatory mediators such as iNOS, COX-2, TNF-α, and IL-6 through the inhibition of NF-κB/ERK/JNK-related signaling pathways. These findings highlight the potential of tectoridin and tectorigenin to reduce oxidative stress and enhance antioxidant defenses, contributing to neuroprotection against cytotoxicity and apoptosis induced by MPP (Gong et al. 2017).
Additionally, tectorigenin has shown promising results in improving cognitive impairment, hippocampal damage, and reduced myelin density caused by CCI in mice. tectorigenin effectively reversed the changes in levels of IL-1β, TNF-α, TLR4, myeloid differentiation primary response gene 88, p-p65, NLRP3, and ASC in these mice. Moreover, tectorigenin mitigated the effects of OGD/R on apoptosis and TLR4/NF-κB pathway-related factors (Yao et al. 2021). Overexpression of TLR4 counteracted the protective effects of tectorigenin in OGD/R-induced HT-22 cells, indicating that tectorigenin’s role in inhibiting the TLR4/NF-κB signaling pathway is crucial for alleviating cognitive impairment in CCI mice (Yao et al. 2021).
Tectoridin and tectorigenin exhibit remarkable neuroprotective properties, acting through pathways such as PI3K/AKT, PPARγ/NF-κB, and TLR4/NF-κB. These compounds mitigate oxidative stress, regulate mitochondrial function, and inhibit neuronal apoptosis by modulating key proteins, including p-AKT, Bcl-2/Bax, and HIF-1α. Their roles in reducing inflammatory mediators and oxidative damage in Parkinson’s disease models and cognitive impairment highlight their therapeutic potential. Furthermore, tectorigenin’s ability to reverse hippocampal damage and protect myelin integrity underscores its significance in addressing central nervous system injuries and neurodegenerative disorders. These findings offer promising avenues for future neuroprotective therapies.
Cytotoxic Mechanisms and Antitumor Activity
Over the past decade, tectorigenin and tectoridin have been recognized as potent cytotoxic and antitumor agents effective against various malignancies, including liver, colon, lung, breast, gastric, skin, kidney, ovarian, prostate, laryngeal, pancreatic cancers, and leukemia. The cytotoxic effects of tectorigenin and tectoridin in cancer cells and their antitumor activity primarily stem from modulation of key signaling molecules, including G protein-coupled receptor 30 (GPR30), ERK, protein kinase C (PKC), MAPK, matrix metallopeptidase 1 (MMP1), matrix metallopeptidase 2 (MMP2), caspase3, PI3K, AKT, NF-κB, signal transducer and activator of transcription 3 (STAT3), TNF-α, COX-2, G1/S-specific cyclin-D1 (cyclin D1), cyclin-dependent kinase (CDK), matrix metallopeptidase 9 (MMP-9), baculoviral IAP repeat-containing 5 (BIRC5), cellular inhibitor of apoptosis protein 1 (cIAP-1), X-linked inhibitor of apoptosis protein (XIAP), Bcl-2, and various caspases.
Breast Cancer
Research suggests that tectoridin exerts its estrogenic effects mainly through the GPR30 and ERK-mediated rapid nongenomic estrogen signaling pathway in Michigan cancer foundation-7 MCF-7 human breast cancer cells. Tectoridin promotes the phosphorylation of ERK1/2 and enhances the accumulation of cyclic adenosine monophosphate (cAMP) within the cells, a key feature of GPR30-mediated estrogen signaling (Kang et al. 2009). Zeng et al. (2018) reported that tectorigenin effectively inhibits the proliferation of MDA-MB-231 and MCF-7 breast cancer cells in both a dose- and time-dependent manner. Tectorigenin induces apoptosis and causes G0/G1-phase cell cycle arrest while also suppressing cell migration and invasion. Treatment with tectorigenin reduces the expression of matrix metalloproteinases MMP-2, MMP9, BCL-2, phosphorylated-AKT, and MAPK signaling pathways. At the same time, tectorigenin increases the expression of Bax, cleaved poly polymerase, and cleaved caspase-3. By downregulating the AKT and MAPK signaling pathways and upregulating the expression and activity of the caspase family, tectorigenin shows promise as a potential therapeutic agent for human breast cancer (Zeng et al. 2018).
Colon Cancer
Tectoridin has been shown to suppress the proliferation, migration, and invasion of colon cancer cells (Caco-2 cells) in a dose-dependent manner while promoting apoptosis. Additionally, tectoridin dose-dependently reduced the expression of PKC and MAPK. Importantly, when the PKC pathway was inhibited, the antitumor effects of tectoridin were nearly abolished, highlighting the critical role of this pathway in tectoridin’s action (Xiong et al. 2021). Tectorigenin also demonstrated the ability to impede the invasion of human Caco-2 cells (Dai et al. 2015; Wani et al. 2017). In a study by Dai et al. (2015), reverse transcription-polymerase chain reaction (RT-qPCR), and enzyme-linked immunosorbent assays were used to explore how tectorigenin inhibits TNF-α-induced C-X-C motif chemokine ligand 10 (CXCL-10) expression in Caco-2 cell line. Tectorigenin inhibits C-X-C motif chemokine ligand 10 (CXCL10) production by blocking TNF-α-mediated NF-κB activity.
Prostate Cancer
Wang et al. (2013b) reported that tectorigenin exhibits the most potent activity, with an IC50 value of 0.08 μM. Tectorigenin showed significantly stronger cytotoxic activity effects in RM-1 prostate cancer cells compared to its glycoside, tectoridin.
Lung Cancer
Subcutaneous administration of tectorigenin at 30 mg/kg over a 20-day regimen in murine Lewis lung carcinoma (LLC)-implanted mice elicited a significant 30.8% reduction in tumor volume. Concurrently, intraperitoneal delivery of tectorigenin and tectoridin at the same dosage for 10 days in Institute of Cancer Research (ICR) mice bearing sarcoma 180 (S180 cell line) tumors demonstrated marked suppression of tumor mass, with reductions of 44.2 and 24.8%, respectively (Jung et al. 2003). Tectorigenin also demonstrated a dose-dependent inhibition of cancer growth in adenocarcinomic human alveolar basal epithelial cells (A549) cells and caused a dose-dependent decrease in Kristen rat sarcoma viral oncogene homolog (KRAS) expression, further supporting its cytotoxic activity (Wani et al. 2017). Tectoridin also showed the strongest correlation with Polo-like kinase 1 (PLK1) (Wang et al. 2023). When combined with a PLK1 inhibitor, tectoridin synergistically suppressed autophagy and ferroptosis while promoting caspase-3-mediated apoptosis in A549 cells (Wang et al. 2023). These findings by Wang et al. (2023) highlight a promising drug target and a potential combination therapy strategy involving a PLK1 inhibitor and tectoridin for treating lung adenocarcinoma patients.
Blood Cancer (Leukemia)
Tectorigenin has been demonstrated to induce differentiation of human promyelocytic leukemia human promyelocytic leukemia cell line (HL-60 cells) into granulocytic and monocyte-macrophage lineages, eliciting apoptotic DNA modifications analogous to those observed with genistein. Furthermore, tectorigenin attenuated epidermal growth factor (EGF)-dependent autophosphorylation of the EGF receptor and downregulated Bcl-2 protein expression, albeit with diminished potency relative to genistein. These findings collectively highlight tectorigenin’s potential as a therapeutic candidate for leukemia (Lee et al. 2001). In a separate study, Guo et al. (2016) observed that tectorigenin suppressed osteosarcoma (OS) cell proliferation in a dose- and time-dependent manner Notably, tectorigenin significantly inhibited human bone osteosarcoma epithelial (OS) cell migration and invasion (p < 0.05), concomitant with upregulation of cleaved caspase-3 and suppression of MMP1, MMP2, and MMP9 expression, further substantiating its cytotoxic activity in OS cells (Guo et al. 2016).
Ovarian Cancer
Tectorigenin has emerged as a potential chemosensitizer for paclitaxel-resistant ovarian cancer cells via modulation of the AKT/inhibitor of nuclear factor kappa B (IκB)/inhibitor of IκB kinase (IKK)/NF-κB signaling axis, proposing a novel strategy to overcome chemoresistance in ovarian carcinoma (Yang et al. 2012). Yang et al. (2012) demonstrated that tectorigenin synergistically enhanced paclitaxel-induced growth inhibition in both paclitaxel-resistant (MPSC1 TR, A2780 TR, SKOV3 TR) and non-resistant ovarian cancer models. The tectorigenin-paclitaxel combination triggered caspase-3, caspase-8, and caspase-9-mediated synergistic apoptosis, surpassing monotherapy efficacy (Yang et al. 2012).
Mechanistically, tectorigenin suppressed NF-κB nuclear translocation and downregulated NF-κB-dependent pro-survival mediators, including cellular fas-associated death domain protein-like interleukin-1β-converting enzyme(FLICE)-like inhibitory protein (cFLIP), X-linked inhibitor of apoptosis protein (XIAP), Bcl-2, B-cell lymphoma-extra large (Bcl-xL), and COX-2 (Yang et al. 2012). Concurrently, tectorigenin-paclitaxel co-treatment inhibited IκB/IKK phosphorylation and AKT activation in paclitaxel-resistant cells (Singha et al. 2015). Pharmacological AKT inhibition (LY294002) or dominant-negative AKT overexpression potentiated tectorigenin-paclitaxel-induced growth suppression, whereas constitutively active myristoylated AKT (MYR-AKT) attenuated these effects (Yang et al. 2012). MYR-AKT partially restored NF-κB nuclear translocation and IκB/IKK phosphorylation, underscoring AKT’s regulatory role in tectorigenin-paclitaxel-mediated signaling (Yang et al. 2012).
In prostate cancer models, tectorigenin downregulated prostate derived epithelial factor (PDEF), prostate-specific antigen (PSA), and insulin-like growth factor I receptor (IGF-1R) mRNA expression, concomitant with reduced PSA secretion, IGF-1R protein levels, human telomerase reverse transcriptase (hTERT) transcription, and telomerase activity. Contrastingly, tissue inhibitor of metalloproteinase-3 (TIMP-3) mRNA was upregulated. In vivo, B. chinensis extracts delayed tumor progression in nude mice, implicating tectorigenin’s dual role in apoptosis induction and invasiveness suppression via TIMP-3 upregulation and oncogene downregulation (Thelen et al. 2005).
Tectorigenin induced G0/G1-phase cell cycle arrest in a concentration-dependent manner (200–300 µM), suppressing phosphorylated retinoblastoma protein (p-RB) and cyclin-dependent kinase 4 (CDK4), while elevating p21 expression. Yeh et al. (2020) corroborated these findings in glioblastoma, demonstrating tectorigenin-mediated G0/G1 arrest and proliferation inhibition.
Liver Cancer
Tectorigenin elicited dose- and time-dependent apoptosis in human hepatocellular carcinoma (HepG2) cells, primarily through ROS overproduction, intracellular Ca2⁺ dysregulation, mitochondrial depolarization, cytochrome c release, and caspase-9/caspase-3 activation, rather than non-specific cytotoxicity (Jiang et al. 2012).
Tectorigenin and tectoridin demonstrate significant anti-inflammatory potential by modulating key signaling pathways and reducing inflammatory mediators. Studies reveal their efficacy across diverse models, including their ability to inhibit critical cytokines, enzymes, and pathways such as NF-κB, COX-2, and MAPK. These compounds exhibit protective effects against oxidative stress, apoptosis, and tissue damage, showcasing therapeutic promise. Their low cytotoxicity further supports their potential for clinical application in inflammatory conditions, underscoring their importance as viable candidates for future drug development.
Anti-alcoholism and Hepatoprotective Activity
Orally administered tectoridin has demonstrated hepatoprotective activity and protection against tert-butyl hydroperoxide-induced cytotoxicity in HepG2 cells by inhibiting apoptosis. Tectoridin may act as a prodrug that converts to tectorigenin, which also possesses hepatoprotective properties (Lee et al. 2005).
Zhang et al. (2019) studied the effects of tectoridin on alcohol metabolism in rats and found that tectoridin exhibited anti-alcoholism properties. Oral administration of tectoridin; 75 mg/kg body weight) demonstrated superior ethanol clearance (p < 0.01 vs. tectorigenin: 43.67%), while preserving peroxisome proliferator-activated receptor alpha (PPARα) expression and its downstream targets—medium-chain acyl-CoA dehydrogenase (MCAD), acyl-CoA oxidase (ACO), and cytochrome P450 4 A (CYP4A)—at transcriptional and enzymatic levels. This protective effect against ethanol-induced liver steatosis was primarily due to tectoridin’s modulation of the PPARα pathway and its ability to improve mitochondrial function (Xiong et al. 2021).
Antihyperglycemic Activity
Moon et al. (2006) reported that tectorigenin exhibits enhanced hypoglycemic activity relative to tectoridin. Intraperitoneal tectorigenin administration (5–10 mg/kg, 7 days) in streptozotocin-induced diabetic rats significantly reduced hyperglycemia, total cholesterol, low-density lipoprotein (LDL)-/very low-density lipoprotein (VLDL)-cholesterol, and triacylglycerides. Tectorigenin improved insulin sensitivity and glucose homeostasis in high-fat diet (HFD)/streptozotocin (STZ) models without inducing weight gain, edema, or cardiac hypertrophy, likely via PPARγ/NF-κB pat




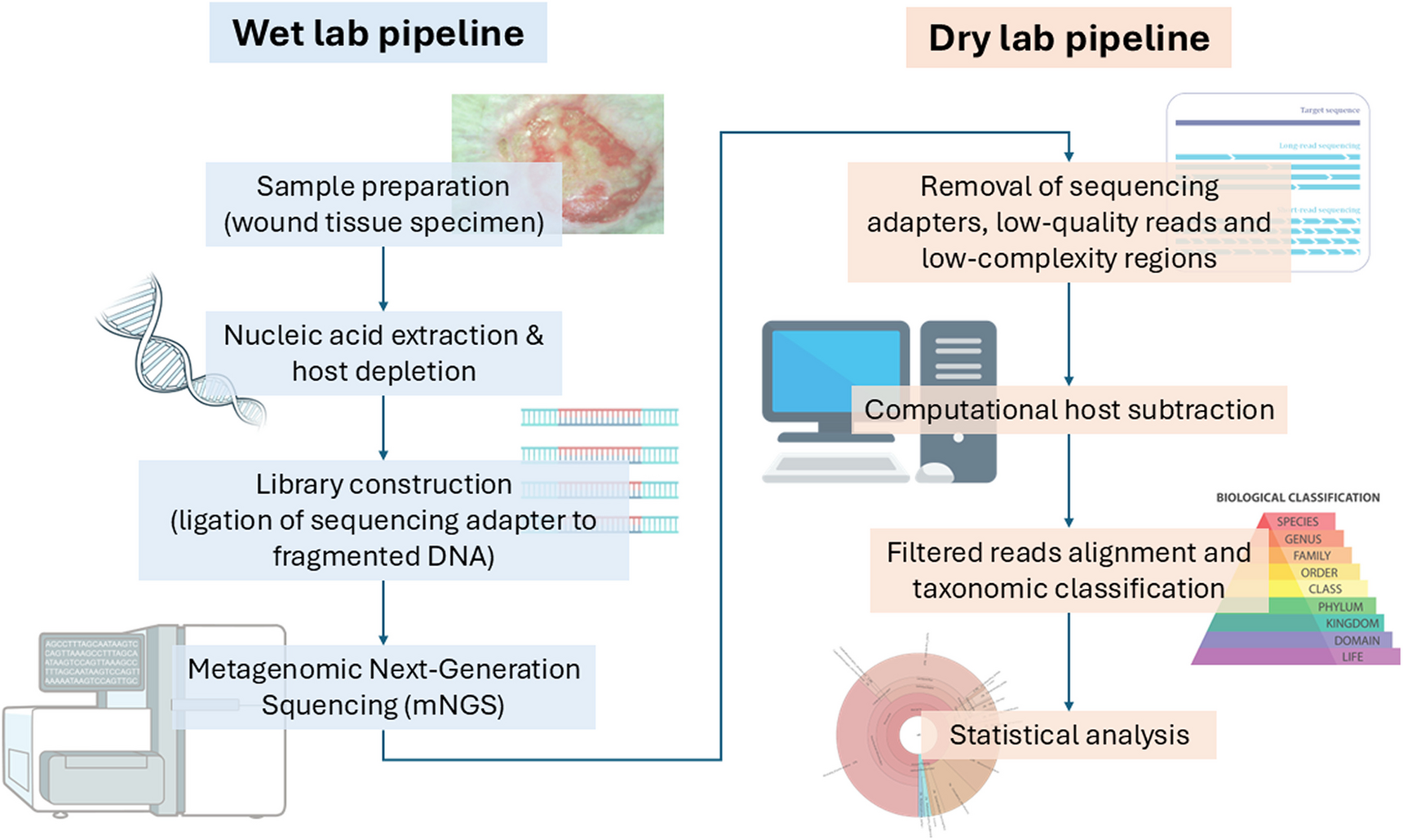

Comments (0)