
Remember me





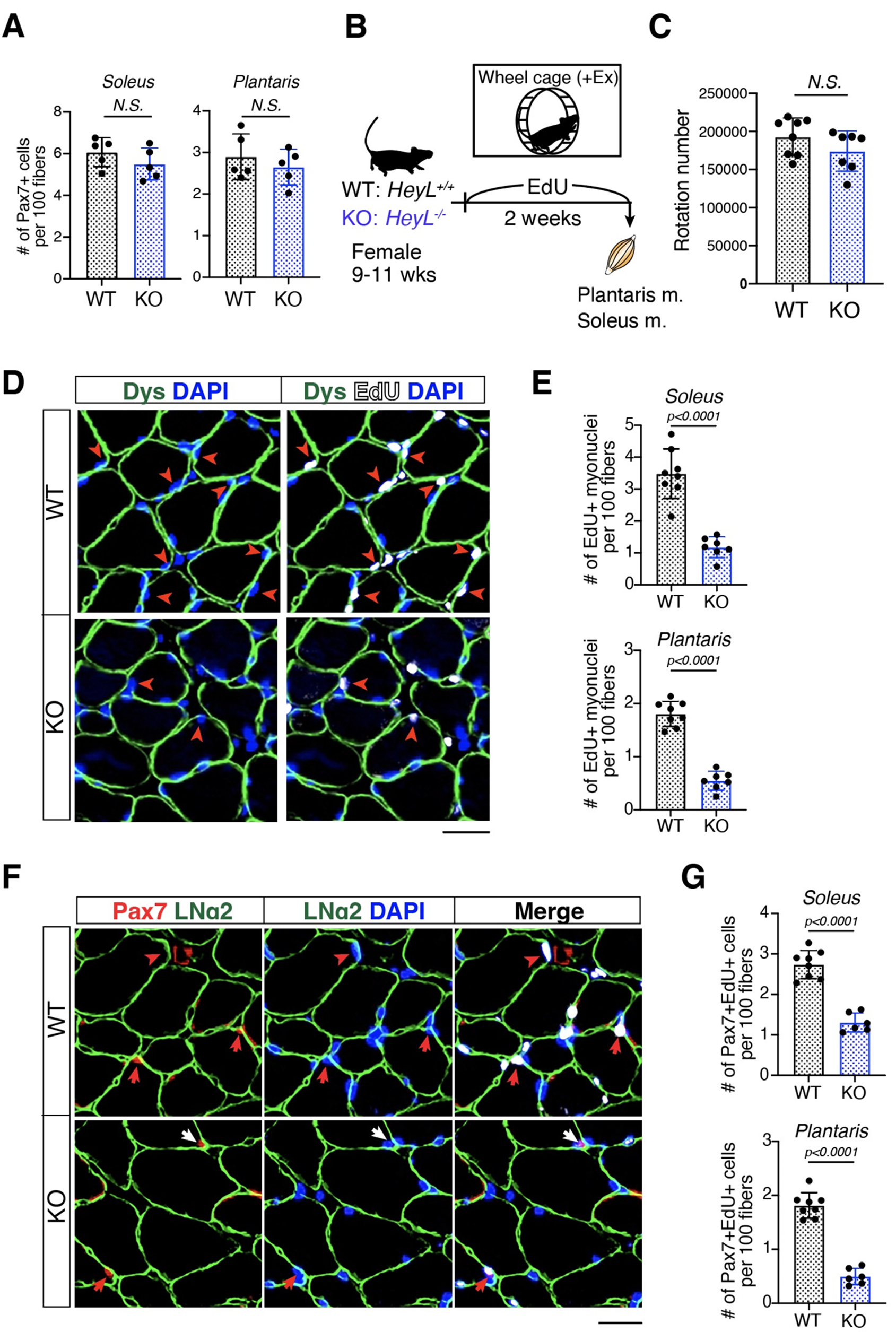
ActRII ligands, including Activin A (ActA), myostatin (or GDF8), and GDF11, have been shown to block myoblast differentiation [13]. We first sought to compare the signaling and functional effects of ActRII ligands versus BMP ligands in human skeletal muscle cell culture.
Primary human muscle cell lines can serve as an in vitro surrogate for how ligands may act in humans [13]. However, primary cultures have several limitations, including restricted proliferation capacity and applicability for stable cell line production. As such, we generated and characterized a human skeletal muscle cell line transduced with human telomerase reverse transcriptase (hTERT) and a temperature-sensitive SV40 large T antigen (tsSV40). In brief, a single cell clone was selected and characterized based on SV40 expression at non-permissive temperatures and molecular profiling was conducted to compare expression of relevant myogenic markers to primary cells at corresponding days of differentiation. These clonal transduced human Skeletal-Muscle Derived Cells (termed iHUSKMDC or iSKMDC in this report) are shown to be myoblasts with an ability to differentiate into myotubes and exhibited similar behavior to primary cells based on molecular profiling (Supplemental Fig. 1A and 1B).
We first compared relative canonical SMAD1/5 signaling across the BMP ligand family and selected ligands representing distinct sub-classes [18] and potencies, including ligands previously linked to muscle growth (Fig. 1A; upper left). Next, we followed up with further experiments to investigate the downstream effects these BMP ligands—BMP7, BMP9, BMP10, or BMP14—compared to ActRII ligands—ActA, GDF8, or GDF11.
Fig. 1
ActRII and BMP ligands promote distinct SMAD signaling yet similar differentiation blockade in human myoblasts. (A) Western blotting analysis for SMAD signaling with BMP ligands at 300 ng/ml (upper left), SMAD signaling with a dose response of ActRII and BMP ligands (bottom left), and non-SMAD signaling with a dose response of ActRII and BMP ligands (right). Human transduced myoblasts (iHUSKMDC) were acutely stimulated with vehicle as a negative control, BMP ligands, or ActRII ligands for 30 min. (B) Western blotting analysis with combined ligands. Human iHUSKMDC myoblasts were stimulated with vehicle as a negative control or with increasing doses of ActA ± BMP9 (left) or ActA ± BMP7 (right) at 1, 10, or 100 ng/ml. (C) Human myoblast differentiation assay with individual ligands. iHUSKMDC myoblasts were differentiated into myotubes with vehicle as negative control, TNFα (30 ng/ml) or IGF1 (10 ng/ml) as positive controls or increasing doses of ActRII or BMP ligands (0.3, 30, or 300 ng/ml). Representative images are shown per treatment, with myotubes (green) identified using anti-MyHC antibody staining and nuclei (blue) using DAPI staining. Scale bar, 100 μm. Differentiation was quantified by evaluating the percentage of nuclei within myotubes that were positively identified using anti-MyHC antibody staining. Differences between groups were analyzed using one-way ANOVA (compared to UNT group). Data are means ± SD. Asterisks indicate differences with the control. Statistical analyses were conducted with a One-way ANOVA. *p ≤.05; **p ≤.01; ***p ≤.001; ****p ≤.0001 compared to control
Consistent with previous results in other cell types, in human skeletal muscle cells, ActA, GDF8, and GDF11 acutely induce SMAD2 phosphorylation whereas BMP7, BMP9, BMP10, and BMP14 promote SMAD1/5 phosphorylation in a dose-dependent manner (Fig. 1A; bottom left). Among the BMP ligands tested, BMP9 and BMP10 showed stronger potency compared to BMP7, with BMP14 as the relatively weakest activator of SMAD1/5 signaling.
Given that prior reports have suggested involvement of non-SMAD signaling within the TGFβ superfamily, we next sought to determine the effects of ligands on alternative pathways [27,28,29]. Of note, it was previously suggested that BMPs may promote positive skeletal muscle regulation via the pro-hypertrophy AKT/S6 pathway [20, 21]. ActRII ligands (GDF8 and ActA) and BMP ligands (BMP7 and BMP9) acutely induced their respective SMAD signaling in fully differentiated myotubes without strong induction of p38 or AKT/S6 signaling (Fig. 1A; right). To determine wither ligands may have longer term effects, we also treated cells for 24-hours and failed to observe consistent BMP-mediated increased pAKT or pS6 in myoblasts or myotubes despite previous studies suggesting activation of this pathway [20, 21] (Supplemental Fig. 1C).
Multiple reports have suggested competition and interplay between the canonical ActRII and BMP axes, both at the receptor and SMAD signaling levels [20, 30, 31]. As such, we selected ActA, BMP7, and BMP9 to further test a dose response of co-treatment in human myoblasts and assess changes in their downstream SMAD response. Combined ActA and either BMP showed minimal, if any, acute crosstalk inhibition of their respective phosphorylated SMAD signaling at the doses tested. (Fig. 1B).
Finally, to determine their functional effects on myogenic differentiation, human myoblasts were allowed to differentiate for 5 days in the presence or absence of ligand (0.3–300 ng/ml) and we assessed differentiation by myosin heavy chain (MYHC) staining. Doses were chosen based on the observed phosphorylated SMAD signaling patterns. As positive controls, the proinflammatory cytokine TNFα blocked differentiation, whereas the pro-anabolic IGF1 promoted differentiation (Fig. 1C). In line with prior reports [4, 13], ActRII ligands significantly inhibited differentiation in a dose-dependent manner. All BMP ligands tested blocked differentiation to varying degrees relative to their phosphorylated SMAD1/5 potencies, confirming a common myogenic functional effect across these TGFβ family members. Taken together, in human muscle cells, ActRII and BMP ligands promote distinct canonical SMAD responses with minimal signaling crosstalk yet show similar blockade of myogenic differentiation.
Receptor-activated SMAD (R-SMAD) overexpression promotes blockade of human myoblast differentiationAlthough non-SMAD pathways have been previously reported for TGFβ family members, 25–27 in our human muscle cells, we failed to see clear, consistent alternative signaling. As such, for the first time, we generated SMAD-overexpressing skeletal muscle cell lines to assess the direct effects of individual SMADs within this cell context. Specifically, we generated stable overexpression cell pools of wild-type receptor activated SMADs or R-SMADs—SMAD1, SMAD5, SMAD2, or SMAD3. The empty vector control line was able to differentiate comparable to the parental iHUSKMDC cells (Supplemental Fig. 2A). Total protein levels of each SMAD were validated by western blotting analysis. Overexpression of each R-SMAD was associated with increased corresponding phosphorylated R-SMAD at baseline, demonstrating induction of SMAD signaling in the absence of exogenous ligand (Fig. 2A and B). Persistent SMAD signaling was observed in each cell line throughout the myogenic differentiation process (Supplemental Fig. 2B.) Expression profiling of this skeletal muscle cell system showed detectable levels of endogenous ActRII and BMP ligands, suggesting an autocrine feedback response (Supplemental Fig. 1B).
Fig. 2
R-SMAD overexpression blocks human myoblast differentiation and ActRII inhibition mitigates ligand inhibitory effects. (A) Differentiation assay and western blotting analysis validation for SMAD2 and SMAD3 over-expressing lines. Stable skeletal muscle cell lines (iHUSKMDC) with SMAD2 or SMAD3 over-expression (OE) were differentiated into myotubes. Representative images (n = 3 replicates) are shown for each treatment, using anti-MyHC antibody staining for myotubes (green) and nuclei (blue) using DAPI staining. Scale bar, 100 μm. Western blotting analysis at day 0 of differentiation was used to confirm overexpression and signaling with SMAD-selective antibodies. (B) Differentiation assay and western blotting analysis validation for SMAD1 and SMAD5 over-expressing lines. Stable skeletal muscle cell lines (iHUSKMDC) with SMAD1 or SMAD5 over-expression (OE) at lower and higher levels were differentiated into myotubes. Representative images (n = 3 replicates) are shown for each treatment, using anti-MyHC antibody staining for myotubes (green) and nuclei (blue) using DAPI staining. Scale bar, 100 μm. Western blotting analysis at day 0 of differentiation was used to confirm relative overexpression and signaling with SMAD-selective antibodies across the lines. (C) Differentiation assay and western blotting analysis to assess ActRII inhibition effects. Human myoblasts (iHUSKMDC) were pre-treated with a selective anti-ActRIIA/B neutralizing antibody or isotype control (25 or 100 nM) followed by stimulation with 10 ng/ml ActA, BMP7, or BMP9. Scale bar, 100 μm. Representative images (n = 2) are shown per treatment, with myotubes (green) identified using anti-MyHC antibody staining and nuclei (blue) using DAPI staining. Scale bar, 100 μm
To assess functional implications of R-SMAD overexpression, myoblasts were allowed to differentiate for 5 days without added recombinant protein ligands that activated SMAD signaling (ActRII ligands and BMPs), followed by anti-MYHC staining. Overexpression of SMAD2 or SMAD3 alone blocked myoblast differentiation into myotubes, similar to exogenous ActRII ligand treatment and in line with prior reports [4, 13], demonstrating that SMAD2 or SMAD3 signaling is sufficient to inhibit myogenic differentiation (Fig. 2A). Overexpression of SMAD1 or SMAD5 blocked myoblast differentiation in a dose-dependent manner, with higher SMAD1/5 signaling corresponding to the inhibitory phenotype, suggesting a SMAD signaling threshold effect (Fig. 2B). Together these data indicate that R-SMAD signaling through either the SMAD2/3 or SMAD1/5 axes mediates the inhibitory effects of ActRII and BMP ligands on myogenic differentiation.
ActRII inhibition rescues activin A and BMP ligand-mediated effects on human skeletal muscle cellsReceptor binding and signaling by ActRII and BMP ligands are highly promiscuous, given that ligands across sub-families have been shown to bind and engage with activin type II receptors (ActRIIA and ActRIIB) and the BMP type II receptor (BMPR2) [32, 33]. Multiple BMP ligands, including BMP7 and BMP9, can reportedly bind and signal through ActRII receptors within some cell types [34, 35]. Previous studies demonstrated that BMP7 can bind with high affinity to ActRIIA relative to BMPR2 whereas BMP9 has a relatively higher affinity to BMPR2 [33, 36].
Prior publications have demonstrated that antibody-mediated ActRIIA/B inhibition (αActRII) is sufficient to reduce ActA and GDF8 downstream SMAD signaling as well as attenuate their blockade of skeletal muscle differentiation [16]. Whether ActRII signaling is functionally relevant to BMP ligands within skeletal muscle cells, however, is unclear. As such, myoblasts were treated with ActA, BMP7, or BMP9 in the presence or absence of an αActRII antibody, either acutely for signaling or for multiple days to assess myogenic differentiation. Consistent with previous results, αActRII blocked ActA-mediated SMAD2 phosphorylation and rescued myogenic differentiation [13, 17] (Fig. 2C). Surprisingly, both BMP7- and BMP9-mediated signaling and differentiation blockade were at least partially rescued with the αActRII antibody. BMP7 was more responsive to antibody treatment, showing similar rescue compared to the canonical ActRII ligand, ActA; this is notable since these cells also express BMPR2, which is not bound by the anti-ActRIIA/B antibody and, thus, amenable for ligand binding (data not shown) (Fig. 2C). Similarly, αActRII-mediated blockade of SMAD1/5 signaling was also observed with other members of the larger BMP ligand family (Supplemental Fig. 2C). Thus, in skeletal muscle cells, BMP ligands can promote downstream effects in a partly ActRII-dependent fashion, demonstrating a putative key role for BMP inhibition with the use of anti-ActRIIA/B neutralizing antibodies.
ActRII and BMP ligands suppress myogenic markers in differentiating human myoblasts and differentiated human myotubesSince we found that both ActRII ligands and BMPs functionally inhibit human myoblast differentiation into myotubes, we next sought to assess the effects of select representative ligands on the expression of key myogenesis markers. Treatment of differentiating (day 0 of differentiation) iHUSKMDC myoblasts with ActA, BMP7, and BMP9 for 24 h suppressed mRNA levels for the myogenic marker myogenin (MYOG), as well as the mRNA levels for myosin heavy chain (MYHC) genes (MYH1, MYH2, MYH3, MYH8) (Fig. 3A). Prior data demonstrated that BMP signaling can have pro-osteoinductive effects on multiple cell types, including resident Sca1+ muscle stromal cells [37]. To determine whether BMPs might have an early osteo-inductive influence on myoblasts, expression of osteogenic differentiation markers were also assessed. Sclerostin (SOST), a negative regulator of bone formation, was significantly increased in response to BMP7 and BMP9; however, expression of the osteoblast marker osteocalcin (BGALP) was significantly repressed and RUNX2, an early osteogenic differentiation marker, was unchanged (Supplemental Fig. 3).
Fig. 3
ActRII and BMP ligands similarly repress myogenic markers in human myoblasts and differentiated human myotubes. (A) qPCR analysis of myogenic differentiation gene markers in ligand-treated myoblasts. Human myoblasts (iHUSKMDC) were stimulated with vehicle as a negative control or single 300 ng/ml dose of ActA, BMP7, or BMP9 for 24 h with downstream gene expression determined by qPCR (n = 4/treatment). Data are mean ± standard deviation (SD) from the mean. Asterisks indicate differences with the control. Statistical analyses were conducted with a One-way ANOVA. *p ≤.05; **p ≤.01; ***p ≤.001; ****p ≤.0001 compared to control. (B) Heatmap for muscle and bone marker gene expression profiles in ligand-treated human myotubes. ActRII and BMP ligands show similar transcriptional signature on myogenic markers (left). Primary human myotubes (HUSKMDC) stimulated as above were analyzed by qPCR for select genes (n = 5/treatment) (right). Data are mean ± standard deviation (SD) from the mean. Asterisks in the heatmap indicate significant differential expression between the control and the treatment. Asterisks in the bar plot indicate differences with the control group. Statistical analyses were conducted with a One-way ANOVA. *p ≤.05; **p ≤.01; ***p ≤.001; ****p ≤.0001 compared to control
To further elucidate the transcriptional networks associated with ActRII- and BMP-ligand-mediated responses in differentiated muscle cells (day 8 of differentiation), we performed RNA sequencing using single or combined ligand treatment across multiple ligands in each family. Primary human HUSKMDC myotubes were treated for 24-hours with ActA, GDF8, GDF11, BMP7, BMP9, BMP14 or combined ActA with BMP7, BMP9, or BMP14 to confirm our data in myoblasts. Genes were considered differentially expressed if their expression was perturbed > 1.5-fold compared to vehicle with a Benjamini-Hochberg adjusted p value < 0.05.
In differentiated myotubes, pro-atrophy marker MAFBX expression was notably unperturbed whereas MURF1 expression was significantly reduced with BMP7 or BMP9 and trended toward a decrease with all other ligands (Fig. 3B; left). However, all ligands significantly repressed MYOD1 as compared to baseline, except for BMP14, which showed a downward trend. A similar trend was observed with MYOG across ligand treatment. Subsequent qPCR validated these biomarkers, showing a significant repression of both MYOD1 and MYOG with all ligands, along with decreased expression of MYH2 (Fig. 3B; right)—similar perturbations of myogenic marker in myoblasts and consistent with prior reports on the mechanism underlying the negative regulation of human muscle cells by GDF8 [38].
Hepatic over-expression of BMP9 in mice results in liver toxicity with a cachexia-like phenotype and an induction of a pro-atrophy gene program in skeletal musclesBMP9 promotes heterotopic ossification of isolated mouse muscle resident Sca1+ stromal cells ex vivo [37] and was recently shown to block isolated primary murine skeletal muscle cell differentiation, but its full effects on adult skeletal muscle remain unclear [22]. Given our findings in cultured iHUSKMDCs, we investigated the effects of hepatic over-expression of BMP9 in mice. We delivered 20 ug of murine BMP9 DNA plasmid into C57BL6J male mice by hydrodynamic tail vein injection. The control group received an empty DNA vector. Hydrodynamic delivery (HDD) of BMP9 plasmid induced hepatic overexpression of the BMP9 gene, Gdf2 (Supplemental Fig. 4A). Seven days following BMP9 over-expression, mice lost ∼ 20% body weight and displayed a reduction in blood glucose levels (Fig. 4A), which is consistent with prior reports on BMP9-mediated glucose metabolism [39, 40]. Of note, BMP9 HDD increased liver damage enzymes: alanine aminotransferase (ALT) and aspartate aminotransferase (AST) (Fig. 4A), suggesting that local BMP9 over-expression was associated with hepatotoxicity that could have had systemic effects. Weights of heart, liver, and epididymal fat were lower in BMP9 over-expressing mice compared to controls by ∼ 22%, ∼ 27%, and ∼ 35% respectively (Fig. 4B), demonstrating a global wasting syndrome.
Fig. 4
BMP9 overexpression promotes liver toxicity, multi-organ wasting, and elevated muscle atrophy markers despite BMP-SMAD1/5 signaling. (A) Body weight, blood glucose, and levels of liver damage enzymes: alanine aminotransferase (ALT) and aspartate aminotransferase (AST). (B) Heart, liver and epididymal fat pat weights. (C) Weights of lower limb skeletal muscles: quadriceps, gastrocnemius, tibialis anterior, and soleus. In all graphs, light gray bars represent a control group that received an empty vector via a hydrodynamic tail vein injection (n = 11–12) and dark gray bars represent a group that received 20 ug of BMP9 DNA (n = 14). (D) Impact of hepatic BMP9 over-expression for 7 days on signaling in quadriceps muscles. Immunoblots for phosphorylated SMAD1/5 and phosphorylated SMAD2 and total SMAD1 and total SMAD2/3; phosphorylated ribosomal protein 6 S and total S6, MuRF1 and MAFbx in quadriceps muscles of mice that received DNA with an empty vector (control) or BMP9 DNA (BMP9). Immunoblots for vinculin as shown as protein loading controls. Molecular weights are shown on the left-hand side for each blot and arrows point to the protein bands of interest. (E) Impact of hepatic BMP9 over-expression for 7 days on gene expression in quadriceps and gastrocnemius muscles. mRNA amounts for specific genes were quantified by real-time quantitative polymerase chain reaction and expression levels were normalized to a geometric mean of reference genes. Vertical dotted lines in each graph show mRNA levels for each gene in the control group that received DNA with an empty vector (n = 12). Dark gray bars show mRNA levels for specific genes in the group that received BMP9 DNA (n = 14), relative to the control group. Data are mean ± standard deviation (SD) from the mean. Statistical analyses were conducted with an un-paired t-test. *p ≤.05; **p ≤.01; ***p ≤.001; ****p ≤.0001 compared to control
Hepatic BMP9 over-expression resulted in skeletal muscle atrophy with quadriceps muscle weights lower in BMP9 over-expressing mice compared to controls by ∼ 13% (Fig. 4C). Weights of the gastrocnemius, the soleus, and tibialis anterior muscles were not significantly affected at the time of takedown (Fig. 4C). To determine the impact of high levels of BMP9 on the regulation of protein synthesis and degradation markers in skeletal muscles, we performed immunoblotting in quadriceps because this muscle showed the weight loss (Fig. 4D). Compared to control animals, in BMP9 over-expressing mice, phosphorylation of SMAD1/5 was higher, while phosphorylation of SMAD2 was lower in quadriceps muscles (Fig. 4D), demonstrating on-target BMP effects in muscle without compensatory SMAD2 pathway activation. Levels of phosphorylated as well as total ribosomal protein S6, a downstream target of the mTORC1/S6K1 protein synthesis pathway, were lower in BMP9 overexpressing mice compared to a control group by ∼ 82% and ∼ 30% respectively (Fig. 4D). We also quantified protein amounts for the E3 ubiquitin ligases MuRF1 and MAFbx, which are transcriptionally upregulated under atrophic conditions, and induce muscle atrophy via proteasomal protein degradation pathway 31. MuRF1 and MAFbx protein levels were higher in quadriceps muscles of BMP9 over-expressing mice compared to controls by 1878% and 374.5% respectively and correlated with SMAD1/5 phosphorylation levels (Fig. 4D; Supplemental Fig. 4B), demonstrating a pro-atrophic condition.
We next performed gene expression analyses in two muscles: quadriceps, which atrophied with BMP9 over-expression, and gastrocnemius, which did not atrophy. Both muscles showed a similar mRNA expression profile with up-regulation of pro-atrophy markers MAFbx/Fbxo32, MuRF1/Trim6331 along with the metallothioneins Mt1 and Mt232,33 (Fig. 4E). Of the canonical SMAD2/3 activating myokines, only Mstn (or Gdf8) levels were increased in BMP9 over-expressing mice. In line with the myogenic biomarkers observed in cell culture, we observed reduced expression of Myod and Myog as well as the myosin Myh2 with BMP9 over-expressing mice, further supporting a direct on-target BMP9 effect in skeletal muscle (Fig. 4E). Thus, despite differences in observed muscle mass effects, both gastrocnemius and quadricep muscles showed a near identical pro-atrophy gene signature with a concomitant suppression of myogenic genes and myosins, suggesting that skeletal muscle is primed for wasting in the setting of elevated systemic BMP9 levels.
Taken together, despite the increased SMAD1/5 phosphorylation observed, there were increased levels of Mt1 and Mt2 along with Foxo1, MAFbx/Fbxo32 and MuRF1/Trim63, as well as reduced levels of phosphorylated S6, indicating decreased mTORC1/S6K1 signaling in a pro-atrophic state in the absence of SMAD2/3 signaling. In addition, SMAD1/5 phosphorylation levels correlated with elevated atrophy signaling and repressed Myh2 expression (Supplemental Fig. 4B). Therefore, our results contrast prior models showing that elevated BMP-SMAD1/5 signaling is dominant and sufficient to promote a pro-hypertrophy response [19].
Intramuscular over-expression of BMP7 and BMP9 in mice results in heterotopic ossification in skeletal musclesGiven the observed liver toxicity with hepatic BMP9 overexpression, we next assessed the intramuscular overexpression of BMP9 and BMP7 to determine local effects and compare functionalities across ligands. Previously, it was shown that local intramuscular AAV: BMP7 delivery was sufficient to promote skeletal muscle hypertrophy 28 days following injection with increased Igf1 expression and elevated AKT/S6 signaling in wild-type mice. In addition, muscle specific over-expression of BMP7 was shown to have a protective effect in the setting of denervation-induced muscle atrophy19 [20]. We designed rAAV1 vectors encoding either murine BMP9 or BMP7, which were delivered to the tibialis anterior (TA) muscles of adult 13-week-old mice at a similar dose to prior studies to assess chronic 28-day changes. Both AAV1:BMP7 and AAV1:BMP9 increased mRNA expression of their respective BMP ligands by day 7 as evident by RNAscope (Supplemental Fig. 5A). Of note, given our prior observations with hepatic BMP9 by HDD, there was no evidence of hepatoxicity by histology following intramuscular AAV gene delivery at 1 month (Supplemental Fig. 5B). After 1 month, muscle-specific over-expression of both AAV1:BMP7 and AAV1:BMP9 had osteoinductive effects, with a more pronounced phenotype observed with BMP9. AAV1:BMP7 injected mice showed partial ossification of the TA, while AAV1:Bmp9 injected mice had widespread TA ossification (Fig. 5A and B). Thus, at an equivalent AAV dose and treatment time to prior reports, we failed to demonstrate a beneficial effect of muscle, instead observing osteogenic changes.
Fig. 5
Local AAV delivery of BMP7 and BMP9 promotes heterotopic ossification in mouse skeletal muscle. (A) H&E staining of tibialis anterior (TA) cross-sections from mice 1-month post-intramuscular AAV delivery. Representative images of n = 2 mice per group are shown, each at 4X, 10X, and 20X magnification (total numbers of mice were, n = 11 for AAV: BMP7 and n = 12 for AAV1: control or AAV1:BMP9). Boxes indicate select corresponding magnified areas. Arrows indicate areas of ossification. (B) Images of dissected lower limbs from mice1-month post-intramuscular AAV delivery. Representative images of n = 2 mice per group are shown. Arrows indicate areas of ossification
In aged rodent muscles, the TGFβ pathway is upregulated with increased expression of BMP gene markers co-incident with increased pSMAD1/5Previous studies have shown increased SMAD2/3 activation with age in rat muscles [38]. In addition, increased circulating levels of the activin/TGFβ signaling marker follistatin-like 3 (FSTL3/FLRG) have been shown in old human plasma [41]. These observations suggest a regulatory role of activin/TGFβ signaling with age [38, 41]. Of note, BMP7 was among the conserved age-related plasma protein that increased in both humans and mice [41]. Skeletal muscle SMAD1/5 activation was shown to decrease in mice from 1-week to 6-months of age [19], but changes to the pathway in the setting of age-related muscle loss (or sarcopenia) are unclear.
We previously performed comprehensive transcriptional profiling of multiple muscle groups form old rats and identified up-regulated markers of TGFβ signaling co-incident with sarcopenia, suggesting a putative causal role for the pathway in the setting of aging [42, 43]. In these profiled sarcopenic skeletal muscles, we observed consistently elevated BMP markers Id2 and Id3, indicative of an increased BMP signaling axis (Fig. 6A). To confirm changes in BMP signaling with sarcopenia, we assessed SMAD1/5 phosphorylation by immunoblotting in a sarcopenic muscle (tibialis anterior) and a non-sarcopenic muscle (triceps) from rats (Fig. 6B; left). SMAD1/5 phosphorylation was increased in aged rat tibialis anterior, but unperturbed in triceps, demonstrating that the BMP-SMAD1/5 axis is in fact elevated with sarcopenia in rats (Fig. 6B; right).
Fig. 6
Elevated SMAD1/5 signaling and BMP gene targets are observed in aged rat sarcopenic muscle. (A) Heatmap for TGFB pathway gene expression profiles in aged rat diaphragm, gastrocnemius, soleus, and tibialis anterior muscles. Tissues were profiled from multiple time points across the lifespan of rats (from 6 to 27 months). For age-related genes (see the original paper for details), the time points where the genes show significant differential expression vs. 6 months are marked by asterisks. Id1-3, BMP target genes, are highlighted in a black box. (B) SMAD1/5 immunoblotting analysis in select rat sarcopenic and non-sarcopenic muscles. Muscle weights for sarcopenic tibialis anterior (or TA) and non-sarcopenic triceps are shown on the left. Phosphorylated SMAD1/5 and total SMAD1 levels for each muscle group are shown on the right. Data are mean ± standard deviation (SD) from the mean. Statistical analyses were conducted with an un-paired t-test. *p ≤.05; **p ≤.01; ***p ≤.001; ****p ≤.0001 compared to control
Comments (0)