
Remember me



We assembled 213 experimental pairs, comprised of one subject with ASD and one TD control, that were matched, to the best of our ability, for sex, age, self-identified race, and type of dMRI protocol. All pairs were matched for sex (30.0% female). 212 pairs were matched for age; one pair (mean age: 181 months) was separated by 13 months instead of the target, ≤ 12 months. The mean within-pair age difference was 5.2 months. Our subjects ranged from 78–508 months (mean: 182 months). 79% of our pairs were matched for self-identified race. 81% of our subjects self-identified as White, a category that included individuals who identified as Hispanic/Latino. 89% of pairs were matched within the same study and thus were imaged with identical dMRI protocols. A further 4.7% of subjects were paired between studies but were balanced by a pair with the opposite pattern (pair_StudyA-StudyB balanced by pair_StudyB-StudyA). Thus 93.4% of our subjects were matched for dMRI protocol. Within our ASD cohort, 166 subjects (77.9%) were in the ADOS-high category, and 47 subjects (22.1%) were in the ADOS-mid category. We also assembled a distinct cohort of subjects with ADOS scores (rather than relying on the assigned diagnostic category from the originating study) and converted those scores to CSS for comparison across versions and modules of ADOS. This cohort included 160 subjects with a mean CSS of 7.1 (SEM: 0.096; range: 3–10).
The intrastriate location of compartment-like voxelsWe measured the position of each voxel in our matched-volume highly-biased (matrix-like or striosome-like) voxels. The mean location of striosome-like voxels was 0.4 mm more medial (p < 1.0 × 10–100), 5.0 mm more rostral (p < 1.0 × 10–100), and 3.7 mm more ventral (p < 1.0 × 10–100) than the mean location of matrix-like voxels. This matched the expected location biases or striosome established through human immunohistochemistry [54, 55]. Note that since individual voxels were assessed relative to the centroid of either caudate or putamen, the relative positioning of each nucleus within the hemisphere did not drive these differences in location. The locations of matrix-like and striosome-like voxels did not differ, on average, between ASD and TD cohorts. The root-mean-square distance between mean ASD and mean TD voxel location was 0.15 mm for matrix-like voxels, and 0.13 mm for striosome-like voxels.
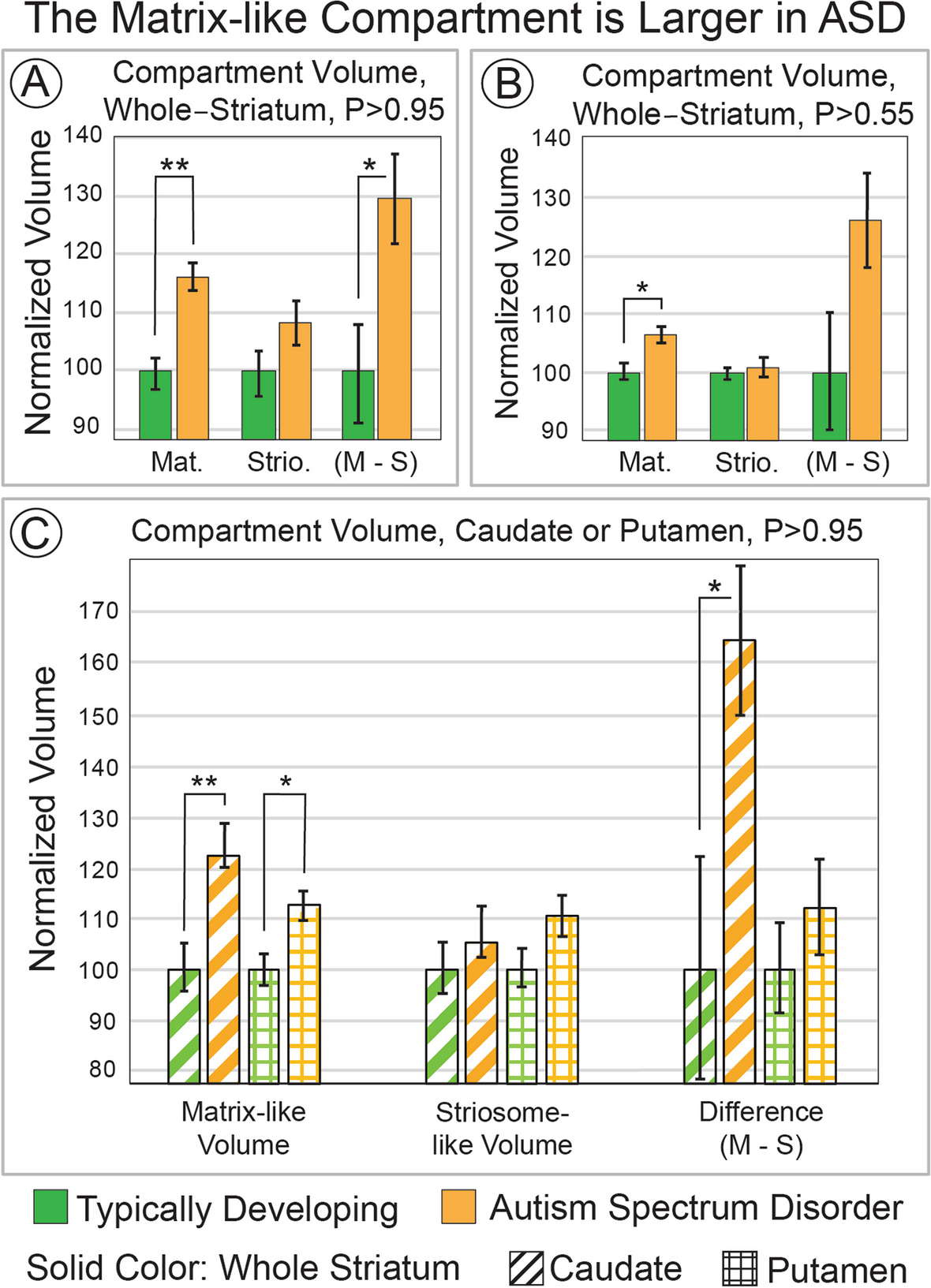
Striatal compartment volume, ASD vs. TDWe extracted the volume of highly-biased voxels (P ≥ 95%, 1.96 standard deviations (SD) above the mean) toward matrix-favoring or striosome-favoring regions (test: hypothesis i). In ASD, the matrix-like compartment was expanded by 16.5% (mean, 245 vs. 211 voxels; p = 9.5 × 10–6; Fig. 1A), while the striosome-like compartment was not significantly different in ASD and TD (mean, 146 vs. 134 voxels; p = 0.10). The within-subject difference in compartment-specific volume (matrix minus striosome) was greater in ASD as well (+ 29.9%; 99 voxels in ASD vs. 76 voxels in TD; p = 0.016). We also assessed compartment-specific volume at P ≥ 0.55, the lowest margin for defining compartment-specific connectivity. Matrix-like volume was expanded in ASD at this minimum probability threshold (mean, 485 vs. 457 voxels, + 6.3%; p ≤ 9.5 × 10–4; Fig. 1B) while striosome-like volume was not different (ASD + 0.71%; p = 0.74). At this minimum threshold, the within-subject differences in.
Fig. 1
The Matrix-like Compartment is Larger in ASD. Striatal voxels that had biased structural connectivity towards matrix-favoring regions (matrix-like voxels) were identified in greater abundance in autism spectrum disorder (ASD) than in matched typically developing controls. This volume expansion was present in highly-biased voxels (panel A, probability threshold ≥ 0.95) and minimally-biased voxels (panel B, probability threshold ≥ 0.55). In panel C, we assessed compartment-specific volume in the two regions that make up the striatum, the caudate and putamen (identified by stripes or checks, respectively). The matrix-like compartment was expanded in both caudate and putamen, but this increase was larger in the caudate. In addition to group-average comparisons, within-individual differences in compartment volume (M–S; matrix volume—striosome volume) suggest that the expansion in matrix-like voxels in ASD is present in individuals with ASD, not solely by group-level differences (A, B, C). Striosome-like voxels did not significantly differ between ASD and controls at any probability threshold, or for any region, even for significance thresholds not adjusted for multiple comparisons (smallest p-value, p ≤ 0.16). **, p ≤ 5.6 × 10–5; *, p ≤ 5.6 × 10–3
compartment volume were larger in ASD, though not a significant degree (mean, 126 vs. 100 voxels, + 26.1%; p = 0.047). The expansion in matrix-like voxels was substantially larger in ADOS-high subjects (+ 19.6%; p = 1.2 × 10–6) than in ADOS-mid subjects (+ 5.3%; p = 0.38).
In a cohort of individuals with complete ADOS data (largely overlapping, but not synonymous with the matched ASD-TD pairs noted above), subjects with elevated CSS (range: 3–10) had elevated matrix-like volume (highly-biased voxels, P ≥ 95%; mean: 248 voxels; SEM: 6.5; Fig. 2). However, increased matrix-like volume did not covary with CSS (coefficient: −3.5; R2 = 0.0027, F(1, 318) = 0.86, p = 0.35, 95% CI [−11.0,4.0]) – the category designation of “autism” was associated with increased matrix-like volume, independent of CSS. That is, group-wise comparisons of matrix-like volume were significantly different but the correlation using continuous variables was not.
Fig. 2
Matrix-like volume vs. ADOS score. In ASD (orange), the Calibrated Severity Score (CSS, which normalizes ADOS scores between versions and modules) had little influence on matrix-like volume. While mean matrix-like volume (P ≥ 0.95) was elevated relative to typically-developing (TD, green) controls, this elevation did not scale with CSS. The dashed line in the elevated ASD box plot (orange) represents the regression coefficient for this cohort (R2 = 0.0027). Note that TD volume measures represent the same mean data shown in Fig. 1. While all subjects were assessed with ADOS, individual scores were provided for few TD subjects. Therefore, TD subjects are illustrated here without CSS scores
Matrix-like volume was larger in ASD in both the caudate and putamen, but the expansion in the caudate was double that found in the putamen (test: hypothesis ii; caudate: + 24.0%, p = 8.2 × 10–5; putamen: + 10.3%, p = 8.1 × 10–3; Fig. 1C). The within-subject difference in compartment volume was significant in the caudate (ASD, + 64.2%; p = 0.017), but not for putamen (ASD, + 11.7%; p = 0.36). Striosome-like volume did not differ between ASD and controls for either caudate or putamen.
The expansion of the matrix-like compartment occurred within a larger striatum. Native-space striatal masks were 3.5% larger in ASD (926 vs. 895 voxels, p = 3.4 × 10–3). This expansion was similar in the left and right hemispheres (+ 3.2% and + 3.9%, respectively). In contrast, the volume of the extra-striate bait regions we used to parcellate the striatum did not differ between ASD and TD (matrix-favoring: + 0.068% in ASD, p = 0.93; striosome-favoring: + 0.32% in ASD, p = 0.73).
Our parcellation technique is based on differential connectivity. Therefore, factors that skew connectivity might lead to an increase in detection of matrix-like voxels in ASD without a true tissue-level expansion in matrix volume. We previously demonstrated that selecting striatal voxels based on FA, rather than selecting for precise connectivity, strongly biases towards a matrix-like pattern of connectivity [58]. We also demonstrated, in healthy subjects, that matrix-like voxels have significantly higher mean FA than striosome-like voxels [58]. Our TD cohort had a highly similar pattern, with FA 4.2% higher in matrix-like voxels than in striosome-like voxels (0.222 vs. 0.213; p = 4.1 × 10–4; Fig. 3). In ASD, FA was 5.4% higher in matrix-like voxels (0.215 vs 0.204; p = 5.5 × 10–6). The ratio of FA between the compartments (matrix-like:striosome-like voxels) did not differ between ASD and TD (p = 0.71). Therefore, it is unlikely that differences in the architecture of matrix-like voxels (as measured by FA) drove increased parcellation of matrix-like voxels. FA in matrix-like voxels did not significantly differ between the cohorts (ASD, −3.2%, NS), while FA in striosome-like voxels was significantly lower in ASD subjects than in TD subjects (ASD, −4.4%, p = 0.015).
Fig. 3
Compartment-specific Fractional Anisotropy. Fractional anisotropy (FA) is higher in matrix-like voxels than in striosome-like voxels in both typically developing (TD) controls and in individuals with autism spectrum disorder (ASD). While FA was significantly lower in the striosome-like voxels of ASD subjects, there was no difference in the FA of matrix-like voxels between TD and ASD. *, p < 0.0378
Identifying a single voxel as matrix-like or striosome-like depends on the relative abundance of streamlines seeded by matrix-favoring vs. striosome-favoring bait regions. We considered the possibility that extra-striate feature of the ASD brain might bias connectivity such that matrix-favoring streamlines were enriched, without a change in the nature of striatal tissue. Increasing the probability that a voxel will parcellate as matrix-like voxels, with no change in the volume of striosome-like voxels, would by necessity pull voxels from the indeterminate pool (0.45 ≤ P ≤ 0.55) into the matrix-like distribution. We found that indeterminate volume did not differ between ASD and TD cohorts (mean of 38.8 vs 41.2 voxels, respectively; p = 0.11), suggesting that the expansion of matrix-like volume in ASD does not result primarily from better tissue parcellation in ASD.
Next, we tested the hypothesis that the increase in matrix-like voxels in ASD is secondary to projections to the striatum, rather than the tissue composition of the striatum. We assessed differences in connectivity in two scenarios. First, we measured the number of completed streamlines following tractography with our extra-striate bait regions as seeds and the whole striatum (not selective for matrix-like or striosome-like voxels) as an obligatory waypoint. Streamlines seeded by both striosome-favoring and matrix-favoring bait regions were reduced in ASD relative to TD (matrix-favoring: ASD, 10.6% less, p = 2.9 × 10–2; striosome-favoring: ASD, 12.1% less, p = 6.4 × 10–3). The ratio of total matrix-favoring:striosome-favoring streamlines within each subject was not significantly different between TD and ASD (p = 0.17). This suggests that the expansion in matrix-like volume we identified was not primarily the result of differences in the structural connectivity of our extra-striate bait regions.
Second, we measured the number of completed streamlines following tractography that set either matrix-like or striosome-like voxels as seeds and all ipsilateral extra-striate gray matter as an obligatory waypoint. In both matrix-seeded and striosome-seeded streamline volumes, fewer streamlines completed tractography in ASD (matrix-like: ASD 9.1% less, p = 4.4 × 10–4; striosome-like: ASD 7.4% less, p = 3.5 × 10–5; Fig. 4). In both ASD and TD, streamlines seeded by striosome-like voxels were more likely to reach gray matter targets than streamlines seeded by matrix-like voxels (ASD: striosome 32.3% more, p = 4.1 × 10–67; TD: striosome 30.3% more, p = 2.6 × 10–72). For individual subjects, striosome-seeded streamlines were more abundant than their matrix-seeded streamlines in 87.1% of TD subjects and 84.0% of ASD subjects. Notably, we previously found this striosome-dominant pattern of connectivity in a different neuroimaging cohort [58]. The matrix-seeded and striosome-seeded mean streamline distributions were highly segregated, with DSCs of 1.9% and 3.1% (ASD and TD, respectively). It is notable that these comparisons of matrix-seeded and striosome-seeded streamlines were identical in their tractography parameters; they utilized matrix-like and striosome-like seed volumes whose volume was identical within each individual and hemisphere, they targeted the same whole-hemisphere gray matter mask, and each iteration of tractography seeded the same number of streamlines per seed voxel. These marked differences in the distribution of streamline bundles between matrix-like and striosome-like voxels did not differ between ASD and TD (p = 0.73).
Fig. 4
Quantitative structural connectivity with compartment-like voxels. In both typically developing (TD) controls and autism spectrum disorder (ASD), striosome-like seed voxels (red) propagate more streamlines than matrix-like seed voxels (blue), despite the fact that seeds are matched for volume and project to identical targets under identical tractographic conditions. However, ASD subjects propagate significantly fewer streamlines than TD subjects for both striosome-like and matrix-like seeds. *, p < 5 × 10–4; **, p < 5 × 10–67
Tissue-level origins of increased matrix-like volumeWe aimed to identify the type of tissue-level change responsible for the expansion in matrix-like volume (Fig. 1). Since matrix-like voxels were defined by differential connectivity, this volume expansion could result from increases in matrix connectivity, reductions in striosome connectivity, or a combination of both changes. As noted above, we found cohort-specific reductions in completed streamlines (ASD vs. TD) but no compartment-specific differences (matrix-like vs. striosome-like) in global measures of structural connectivity in either our bait regions or in our parcellated striatal voxels. This suggested that the ASD-specific changes in matrix-like volume resulted primarily from striatal abnormalities, not to abnormalities in striatal afferents. Three potential tissue-level striatal abnormalities could explain our imaging findings: a decrease in striosome volume, with no change in striosomal architecture (the pattern of striosomal branching); a simplification of the striosomal architecture such that the same volume of striosomal tissue is concentrated in a smaller number of voxels (thicker striosome “tubes,” fewer striosome branches); an expansion in the volume of the matrix with no change in the volume or architecture of the striosome. Though these tissue abnormalities are not mutually exclusive, considering the impact of each abnormality in isolation, and considering how each change would be sampled at the scale of our diffusion voxels, allowed us to predict and assess for three distinct patterns of change in striatal compartment volume (Fig. 5).
Fig. 5
Potential Tissue Alterations Underlying Increased Matrix-like Volume. Each diffusion voxel (resolution = 2 mm isotropic) has the potential to include both matrix and striosome. The character of the tissue included in each voxel determines whether it will be parcellated as matrix-like, striosome-like, or indeterminate. Three potential changes to striatal tissue – a decrease in striosome numbers or projections to the striosome (A); a simplification of striosomal branching and complexity (architecture, B); an increase in matrix volume with no change in striosome (C) – can explain the increase in matrix-like volume we detected in ASD. Granular measurements of compartment bias throughout the probability distribution (histogram analysis, Fig. 6) allow us to infer the nature of the tissue abnormality in ASD that leads to an expansion in matrix-like volume
The striosome is a highly-branched, labyrinthine structure [62] that is largely surrounded by matrix. With diameters of approximately 0.5–1.25 mm in the coronal plane [54, 55], the branches of the human striosome are smaller than our diffusion voxels (2 mm isotropic). Therefore, each striatal voxel may include matrix in isolation or a blend of matrix and striosomal tissue – but never striosome in isolation. At a particular intra-voxel ratio of matrix:striosome tissue (the precise limits of which are the subject of ongoing investigation), that voxel’s connectivity will be indeterminate. On either side of that ratio, connectivity will be biased towards matrix-favoring or striosome-favoring targets. Since voxel position is distributed randomly with respect to striosome location, some voxels will be centered on a striosomal trunk, but many voxels will intersect with branchpoints or cleave the striosome obliquely (Fig. 5, left-most column). While a voxel may reach probability threshold (and thus be parcellated as striosome-like) by including a “direct hit” or by sampling parts of multiple striosomal branches, each of the hypothesized changes to tissue-level composition will shift the likelihood of a direct hit vs. oblique sampling. Each theorized striatal tissue abnormality will therefore produce distinct alterations to the compartment-specific probability distributions.
Within striatal tissue, a reduction in striosome volume, with no change in striosomal architecture (Fig. 5A), would reduce the number of voxels with all types of striosome-like connectivity, both “direct hits” and off-center sampling. With this hypothetical tissue change, all types of striosome sampling – direct hits, oblique cuts, and glancing contacts – would be less common. Therefore, a histogram of connection probabilities would shift toward matrix-like bias throughout the probability distribution. Fewer voxels would include a balanced mix of matrix and striosome tissue, reducing the number of indeterminate voxels.
If striosome volume is unchanged but the architecture of striosome branches is modified (Fig. 5B), different impacts on striatal parcellation emerge. Organizing striosomal volume into fewer, thicker tubules (imagine a net with larger openings but thicker cords connecting each knot) would increase the number of high-probability voxels (since thicker striosome tubules would occupy a larger fraction of “direct hit” voxels). With fewer tubules, fewer voxels would make oblique cuts through the striosome, reducing the number of both low-probability striosome-like voxels and indeterminate voxels. Likewise, thicker, less-abundant striosomal branches would result in a larger number of voxels that included only matrix, as well as a reduction in the volume of obliquely sampled striosome within matrix-like voxels. This hypothetical tissue change would therefore shift volume from the center of the probability distribution to both ends, increasing the volume of both highly biased matrix-like and striosome-like voxels and reducing the volume of low bias and indeterminate voxels.
Increased matrix-like volume could also result from an expansion in the volume of matrix tissue, with no change in the striosome (Fig. 5C). The striosome is surrounded by matrix. Therefore, the number of voxels that sample a mixture of matrix and striosomal tissue is determined by the surface area of the striosomal arborization. If one holds the volume and architecture of the striosome constant, adding additional matrix volume will not increase the amount of overlap between the compartments – adding matrix tissue directly adjacent to a striosome would simply displace other matrix tissue, increasing the number of voxels that sample matrix in isolation. This hypothetical expansion in matrix tissue, with no change in the striosomal compartment, would lead to a selective increase in volume at the uppermost end of the matrix-like probability distribution with no changes in volume at other parts of the probability distribution. Since this increase in matrix tissue would not increase the amount of within-voxel averaging of the striatal compartments, the striosome-like probability distribution, and the volume of indeterminate voxels, would be unchanged.
Histogram analysis of compartment-specific probabilityWe utilized a histogram analysis (test: hypothesis iii; Fig. 6) to differentiate between the tissue-level changes hypothesized above (Fig. 5). We sampled compartment-specific volume along the probability distribution in 0.01-unit increments from 1.0 to 0.55. The expansion in matrix-like volume in ASD occurred entirely in the highest probability (most biased) voxels (Fig. 6); the uppermost three bins contributed 88.0%, 27.8%, and 18.5% of the total volume change, respectively (p = 2.5 × 10–4, p = 0.018, and p = 0.018). The entirety of the expansion in matrix-like volume occurs in the uppermost five bins (the top 11% of the distribution). Striosome-like voxels did not differ significantly between ASD and TD at any point in the probability distribution. As noted above, there was no change in the volume of indeterminate voxels. This pattern of volume changes among the matrix-like, indeterminate, and striosome-like voxels is consistent with an isolated increase in the volume of the matrix compartment in ASD (Fig. 5C).
Fig. 6
Expansion in Matrix-Like Volume Results from a Selective Increase in High-Bias Voxels. Increased matrix-like volume could result from multiple tissue-level changes to matrix or striosome. Histogram analyses can identify the type of voxels (high- vs. low-bias; matrix vs. striosome) that were changed in autism spectrum disorder (ASD) relative to Typically Developing (TD) controls, and thus allowed us to evaluate the potential tissue-level changes hypothesized in Fig. 5. We performed histogram analyses on the matrix-like and striosome-like probability distributions (1.0 to 0.55 for each), quantifying the voxels with compartment-specific connectivity in 0.01 unit bins (each circle = one bin, 45 total bins). Matrix- and striosome-like histograms followed similar patterns, but matrix-like volume diverged in ASD at the highest probability bins (left-to-right = high-to-low probability); below probabilities of approximately 0.95, matrix-like volume was indistinguishable in ASD and TD. Striosome-like volume did not differ between ASD and TD at any part of the probability distribution (data not shown); expansion in matrix-like volume cannot be attributed to a spurious overcount of matrix-like voxels due to a decrease in striosome-favoring connectivity. **, p = 2.5 × 10–4; *, p ≤ 0.02
The influence of experimental factors on matrix-like volume in ASDFour experimental factors influenced the volume of the matrix-like compartment: diagnosis, hemisphere (L/R), striatal volume, and originating study. Sex and self-identified race had no impact on matrix-like volume (p = 0.14 for sex; p-values for race ranged from 0.11–0.39). The regression model including these four significant factors was moderately predictive of matrix-like volume (R2 = 0.32) but highly significant (F(7, 844) = 57.8, p = 1.2 × 10–67). All regression coefficients for these variables were positive. The category of “autism”, as defined by high ADOS scores in the originating studies, correlated with increased matrix-like volume more strongly than the category of “autism spectrum disorders”, as defined by mid-range ADOS scores (t = 3.22 vs. t = 2.46; p = 0.0073 and p = 0.014, respectively), as distinguished by the diagnostic categories assigned by the originating studies. Note that we did not have sufficient ADOS data (measures of individual subjects) to compare matrix-like volume in those designated as “ASD” in the original studies with subjects whose ADOS scores fell in this mid-range. While matrix-like volume was significantly increased in ASD in both the left and right hemisphere, left sided striata were significantly more likely to be increased (left vs. right hemispheres, t = 2.00, p = 0.046). Of these four factors, total striatal volume had the most reliable, but quantitatively the smallest influence on matrix-like volume (t = 15.5, p = 5.6 × 10–7, coeff. = 0.373). Put simply, in most subjects larger striata included a larger number of matrix-like voxels, but this relationship had a smaller impact on inter-individual differences in matrix-like volume than the other three factors.
When assessing the interaction between diagnosis and striatal volume, matrix-like volume was positively correlated with striatal volume in all phenotypic categories (TD, ADOS-mid, and ADOS-high). However, regression coefficients suggested that increasing striatal volume had a slightly greater influence on matrix-like volume in ASD than in TD subjects (ADOS-high: 6.4% larger regression coefficient than TD, t = 16.2, p = 3.0 × 10–4); ADOS-mid: 3.9% larger regression coefficient than TD, t = 13.9, p = 4.0 × 10–4). Study-of-origin significantly impacted matrix-like volume for each study we included. However, the reasons for this impact were impossible to simplify to a single factor: the number of subjects included for a given study and the number of diffusion directions collected by that study had a linear relationship, and p-values for study-of-origin comparisons were therefore inversely correlated with the numbers of both subjects and sampled diffusion directions.
Somatotopic influence on matrix-like volume in ASDWhile one can identify the striosome in every part of the striatum, they are concentrated in the rostral, medial, and ventral striatum [54, 56,57,58]. Layered atop these location gradients, projections from a particular extra-striate region to matrix or striosome are organized somatotopically [56, 58, 62, 63]. We mapped the somatotopic contribution of each of the 10 “bait” regions used to parcellate the striatum (test: hypothesis iv; Fig. 7). Notably, while we mapped left and right hemispheres independently, the somatotopic zones for a given bait region were highly similar between the hemispheres. We extracted the matrix-like volume (P ≥ 0.95) within each somatotopic zone to identify the bait regions with the largest contribution to increased matrix-like volume. In ASD, we found significantly increased matrix-like volume in 7 of 10 somatotopic zones (Table 1, Fig. 8), associated with the following bait regions: PMC, SMC, GPi, anterior insula, posterior orbitofrontal, basal operculum, and basolateral amygdala. Matrix-volume was increased in somatotopic zones specific for both matrix-favoring and striosome-favoring bait regions, and in all parts (rostral:caudal; dorsal:ventral) of both caudate and putamen. We wished to identify whether this bias toward matrix-like connectivity in ASD was limited to the striatum or was also present in the bait regions we utilized for striatal parcellation (test: hypothesis v). We performed quantitative probabilistic tractography between each bait region and the parcellated striatal compartments (equal-volume matrix-like and striosome-like voxels), and then compared ASD and TD on the number of voxels with high bias (P ≥ 0.95) toward matrix-like voxel targets. No bait region had a significant difference between ASD and TD in the bias toward matrix-like voxels, further suggesting that the expansion in matrix-like volume in ASD is driven by abnormalities within the striatum, not from abnormalities in the bait regions used to parcellate the striatum.
Fig. 7
The Somatotopic Organization of Matrix-favoring Projections. Structural connectivity with the human striatum (outlined with light-blue dashes) is organized somatotopically. The five matrix-favoring bait regions utilized here project to all parts of the human striatum (caudate and putamen, indicated by C and P), but at particular zones (colored voxels), matrix-like connectivity is primarily driven by one bait region. Though the left and right hemispheres were parcellated independently, the location of their somatotopic zones was highly similar, as seen in axial (A), coronal (B), and left sagittal (C) planes. No cluster-forming algorithms were utilized. The five striosome-favoring bait regions also have distinct zones of influence, but only the matrix-favoring zones are shown here for clarity. While a somatotopic zone may abut zones specific for other matrix-favoring regions, there was no overlap in somatotopic zones. Somatotopic maps are overlaid on the MNI152_T1_1mm standard brain. Image follows radiographic convention. Coordinates follow MNI convention
Table 1 Normalized matrix-like volume (± standard error of the mean) is expanded in autism spectrum disorder (ASD) relative to matched typically developing (TD) controls in somatotopic zones specific for each of the ten bait regions utilized for striatal parcellation. * indicates a significant increase in matrix-like volumeFig. 8
Regional Contributions to Matrix-like Volume. Mapping the somatotopic zones (seen in Fig. 7) where a given “bait” region had its greatest influence allowed us to quantify the contributions to matrix-like connectivity for each of the 10 bait regions we used to parcellate the striatum. The volume of matrix-like voxels (> 95th percentile) was expanded in ASD for each of the 10 bait regions (orange bars > green bars), though this volume exceeded our corrected significance threshold for only seven regions. Matrix-like volume was expanded throughout the striatum, in somatotopic zones for matrix-favoring regions and for striosome-favoring regions. For each somatotopic zone, volume was normalized to the typically developing (TD) cohort. **, p ≤ 3.0 × 10–3. *, p ≤ 3.0 × 10–2
Comments (0)