
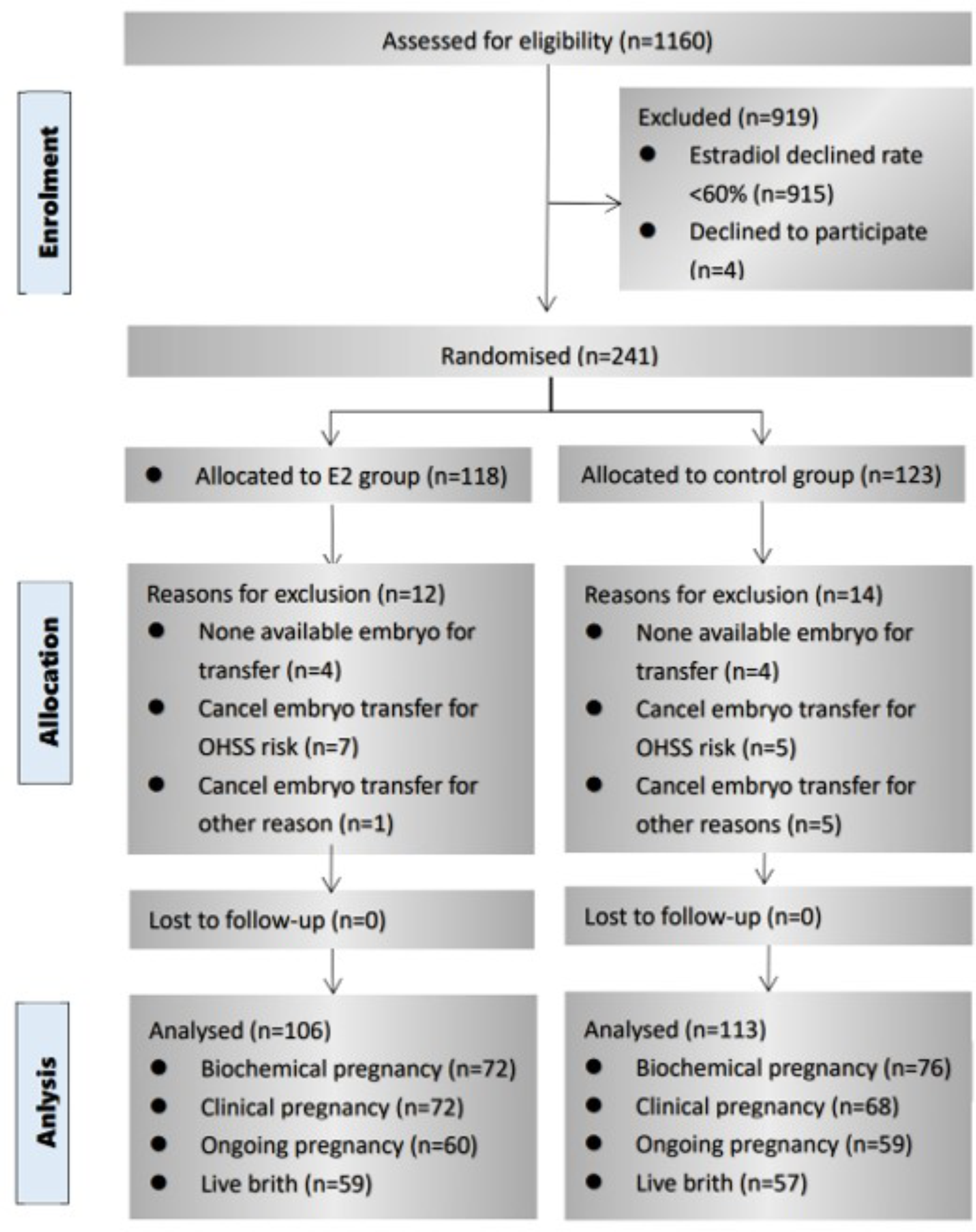
Remember me





In September 2020, a 36-year-old woman was referred to the Reproduction Unit of the Virgen de las Nieves University Hospital of Granada (Spain) due to primary infertility following two years of unprotected sexual intercourse. Patient consent was provided for publication of this report. Her clinical anamnesis includes a family history of endometrial and breast cancer, but no reported cases of infertility. She was a non-smoker with a BMI of 31.5 kg/m². Hormonal analysis revealed a low ovarian reserve (AMH 0.37 ng/mL). Upon vaginal examination, two cervixes of normal size were visualized, separated by a septum in the upper vaginal third. By transvaginal ultrasound, two uteri of normal size were visualized, with normal endometria and ovaries without pathology with a low antral follicle count. The hysterosalpingogram confirmed the uterus didelphys (Fig. 1). The external cervical orifice of the left cervix was cannulated by introducing the catheter into the homolateral uterine cavity and injecting 12 ml of water-soluble iodinated contrast. Filling of the endometrial cavity was observed, which shows no filling defects (Fig. 1A). The left fallopian tube presented dilation of the ampullary portion, with retention of the contrast material, without evidence of passage into the peritoneum, consistent with a non-permeable left hydrosalpinx. Secondly, the external cervical orifice of the right cervix was cannulated, with an injection of 8 ml of water-soluble iodinated contrast (Fig. 1B). Filling of the endometrial cavity was observed, showing no filling defects. The right fallopian tube displayed a normal calibre and appearance, demonstrating the exit of the contrast into the peritoneal cavity (Fig. 1B). A 17 mm cyst with blood-filled content in the right adnexa, suggesting the presence of an endometrioma, was identified using nuclear magnetic resonance imaging (MRI).
Fig. 1
Hysterosalpingogram confirmation of uterus didelphys. A. Cannulation of the external cervical orifice of the left cervix and iodinated contrast revealing filling of the endometrial cavity with no filling defects. The dilation of the ampullary portion of the left fallopian tube, indicative of a non-permeable left hydrosalpinx, is also shown. B. Cannulation of the external cervical orifice of the right cervix and the filling of the endometrial cavity without defects and a normal calibre and appearance of the right fallopian tube with contrast exiting into the peritoneal cavity
Sample collectionAll the samples to characterize the microbiome, metabolome, transcriptome, and immunological features of the uterus didelphys of the recruited patient were collected during the mid-secretory phase of the menstrual cycle (LH +7), measured by LH strips (Clearblue®). For the microbiome analysis, a vaginal swab (V) (eNAT® 606CS01R; COPAN ITALIA, Brescia) and two cervical swabs (eNAT® 606CS01L) (Left cervix, CL; and right cervix, CR) were collected. The collection of endometrial samples from both uterine cavities (Left uterus, UL; right uterus, UR) was carried out using the Tao Brush IUMC endometrial sampler (Cook Medical, Madrid, Spain). To ensure minimal contamination with bacteria from the lower reproductive tract, Tao Brush IUMC was carefully closed within the uterine cavity after sample collection. Subsequently, the samples from the brush were stored in Copan eNAT® transport system (eNAT® 606 C) and stored at a temperature of -80°C.
Subsequently, to study the transcriptome, metabolome, and immunological features of the double uterus, an endometrial biopsy was obtained from both sides (UL; UR) using an endometrial curette device (Gynétics Medical Products, Hamont-Achel, Belgium). The collected tissue was placed into a sterile tube and was divided into three portions. One of them was designated for Pathological Anatomy Service of the hospital to screen for the endometrial pathologies, and to determine the menstrual cycle phase and the presence of CE. Briefly, the first part was fixed in 10% neutral-buffered formalin for conventional histology using haematoxylin-eosin and CD138 (ref. MAD-000735QD-3/V) / MUM1 (ref. MAD-000470QD-3/D) staining following manufacturer recommendation (Vitro Master Diagnostica, Spain). Biopsies were assessed by two pathologists specialized in histological endometrial analysis. The next tissue sample was stored in RNALater for transcriptomic characterization of the endometrial status, and the third biopsy piece was snap-frozen for metabolome analysis. All samples were stored at -80ºC for further analyses.
Endometrial dating and determining chronic endometritisThe histological dating of the endometrium was performed according to the Noyes’ criteria [24]. Both uteri were dated as being in the mid-secretory phase, corresponding to the sampling day, i.e., the cycle day 21. CE was diagnosed through the identification of 1 or more CD138 and/or MUM1 positive plasma cells per 10 high-power field examined in immunohistology staining. Notably, CD138 staining was negative in both endometrial samples (data not presented). The pathologists confirmed the presence of CE in the right-sided uterus, demonstrating MUM1-positive staining for plasma cells, as depicted in Fig. 2B, C and D. Conversely, no MUM1 positive cells were found in the left-sided uterus (Fig. 2A). The concurrent use of CD138 and MUM1 staining mitigates the risk of underestimating the CE diagnosis inherent to single-staining approaches, offering a more reliable assessment of CE [25].
Fig. 2
Histology of the endometrium from both uteri, left (A) and right (B, C and D) stained with MUM1 in immunohistology staining. MUM1-positive cells are indicated by black rings. A and B original magnification X10. C and D original magnification x20
Endometrial receptivity assayed by transcriptomic profileTotal RNA of endometrial tissue samples was extracted using miRNeasy Micro kit (Qiagen, Hilden, Germany) followed by RiboZero kit (Qiagen, Venlo, Netherlands) processing to remove rRNA. The Stranded Total RNA Prep technology (Illumina, USA) was used to generate the libraries. Equimolar libraries were pooled and sequenced using the S2 flow cell, paired-end 100 bp on a NovaSeq 6000 sequencer (Illumina, San Diego, CA, USA). Gene expression profiling of 72 genes analysed with 57 endometrial receptivity-associated biomarkers [14], 11 additional genes relevant to window of implantation (Supplementary Table S1), and four housekeeper genes was estimated using the beREADY algorithm (www.beready.ee, Celvia CC, Competence Centre on Health Technologies, Tartu, Estonia) [26] to confirm the receptivity status of endometrial samples. The results of the beREADY test showed that both endometrial samples, and therefore both uteri, represent early/mid-receptive phase, confirming the endometrial histology dating and demonstrating no differences in endometrial receptivity status between the left and right uteri.
Microbiome analysisThe microbiome of both uterine samples, both cervical samples and vagina were profiled by amplifying the bacterial-specific V4 hypervariable region of the 16S rRNA gene and sequencing. Briefly, DNA was extracted by using the DNA extraction kit (Qiagen QIAamp UCP with Pathogen Lysis Tube S). The primers used were 515F (5’-GTGYCAGCMGCCGCGGTAA) and 806R (5’- GGACTACNVGGGTWTCTAAT). The bioinformatic analysis was performed by using Kraken2 [27]. Microbiome diversity analyses were also conducted under RStudio (R version 4.3.2 (2023-10-31 ucrt)) using phyloseq, vegan, microViz, and ggplot2 R packages [28,29,30]. The relative microbial abundances for the different body sites (i.e., vagina, both cervixes and uteri) are shown in Fig. 3. The vaginal and cervical microbiomes were characterised by clear dominance of lactobacilli. Slight differences in lactobacilli abundance were found between the two cervixes (CL = 96.6% vs. CR = 90.7%). However, in terms of endometrial microbiome, different microbial compositions were noted (Fig. 3). Concretely, the endometrial microbiome from the right was not dominated by lactobacilli (48.2%), but with other microbial genera like Streptococcus, Staphylococcus, Bacillus and Streptococcus comprising a larger percentage of the microbial composition (Fig. 3). These microbial taxa present in UR have been linked to endometrial dysfunction, and particularly to the CE [31, 32]. What is interesting is that while UR presented less ‘favourable’ microenvironment, also same was reflected in the right cervix, both sites presenting less lactobacilli than the left side samples.
Fig. 3
Heatmap of bacteria relative abundance from vagina (V), left cervix (CL), right cervix (CR), left uterus (UL) and right uterus (UR). The maximum size of the circles indicates a relative microbial abundance grater than 1%
The alpha diversity was evaluated by the Shannon, Simpson and Chao1 indexes. As outlined in Table 1, all values indicate that the least diverse niche was the vagina and cervix, followed by the uterus. The right uterus and cervix displayed higher alpha diversity indexes compared to the left, which is consistent with the observed decrease in lactobacilli abundance in the sites of the right cervix/uterus. The diversity indices indicate that ‘unfavourable’ uterine microenvironment is already detected at the cervical sample level. Also, as previously demonstrated, the diversity increased when ascending from the vagina and cervix to the uterus [16].
Table 1 Estimation of alpha diversity indexes of bacterial populationA principal coordinate analysis (PCoA), based on Bray-Curtis distances measured the dissimilarity of microbial community compositions across various sample sites (Fig. 4). The first principal coordinate (MDS1) accounts for 70.5% of the variation. In contrast, the second principal coordinate (MDS2), explaining 21.4% of the variation, does not clearly differentiate between the remaining sites V, CL, CR, UL, and UR. Despite these findings, UR together with CR samples appears to be positioned further from the central cluster of the other female reproductive tract samples (V, CL, UL), suggesting that the microbial community composition of the UR and CR samples are distinct compared to the other sites within the female reproductive tract. The UR and CR samples show a noticeable deviation from the other reproductive tract sites, particularly compared to its contralateral UL/CL samples. This finding reflects a clear lateral asymmetry within the microbial communities between the two uterine cavities of the patient’s uterus didelphys with an effect also on the cervical microenvironment.
Fig. 4
Principal coordinate analysis (PCoA) plot based on Bray-Curtis dissimilarity. Dots are positioned according to the distances between microbial communities in the different samples: vagina (V), left cervix (CL), right cervix (CR), left uterus (UL), and right uterus (UR)
Metabolomic profileThe untargeted metabolomics analysis of both endometrial biopsies (UR and UL) was conducted at Metabolon Inc., Morrisville, NC, USA. This analysis utilized a system consisting of four separate ultrahigh-performance liquid chromatography-tandem mass spectrometry (UPLC–MS/MS) instruments, as previously described [33, 34]. A total of eight hundred sixty-four metabolites from diverse chemical classes were identified from both endometrial tissue samples using the untargeted metabolomic approach. These metabolites encompassed amino acids, lipids, nucleotides, carbohydrates, and xenobiotics, among others. The list of identified compounds is provided in Supplementary Table S2.
Our comprehensive analysis revealed that both uteri primarily share the same metabolomic profile, with 815 common metabolites detected in both uteri (Fig. 5A). However, variations in the peak areas corresponding to individual compounds were observed between the samples (Fig. 5B). Among the metabolites exhibiting the most significant differences between both uteri were metabolites representing coenzyme A and glutathione metabolism, including 3’-dephosphocoenzyme A and 3’-dephospho-CoA-glutathione. Coenzyme A is implicated in various metabolic pathways, such as fatty acid metabolism and the citric acid cycle [35]. CoA-glutathione likely participates in cellular detoxification processes, which are crucial for maintaining cellular homeostasis, particularly in the uterus [36].
Fig. 5
Metabolomic profile of uterus didelphys. A. Venn diagram representing the numbers of metabolites identified in endometrial samples (Left uterus, UL; right uterus, UR). B. Bar chart illustrating the twenty metabolites with the greatest differences (absolute values) in log peak area detected between the samples (blue for right uterus (UR), orange for left uterus (UL)). The right panel evidences the Log10 differences in peak areas in absolutes values among UR and UL. Bars indicate the magnitude of differences for each metabolite.
It is noteworthy to highlight that several key metabolites were exclusively detected in one sample (Fig. 5A), specifically 13 in the right uterus and 36 in the left uterus (Supplementary Table S2). These metabolites were involved in different biochemical pathways, potentially reflecting different uterine microenvironments (Fig. 6).
Fig. 6
The enriched metabolic pathways in the left (A) and right (B) uterus based on the group of metabolites that were uniquely detected in each uterus. This dotplot chart was done using the MetaboAnalyst 6.0 tool (https://www.metaboanalyst.ca). The enrichment ratio was calculated as the number of hits within a particular metabolic pathway divided by the expected number of hits. P-values were obtained from the Student’s t test univariate analysis after linear regression and pareto scaling normalization
Comments (0)